Recent advances in understanding the terminal Ediacaran Earth-life system in South China and Arctic Siberia
Avances recientes en la comprensión del sistema de vida terrestre del Ediacárico tardío en China meridional y el Ártico siberiano
H. Cui1,2, A.J. Kaufman3, S. Xiao4, D.V. Grazhdankin5,6, S. Peek7, A.J. Martin8, N.V. Bykova5, V.I. Rogov5, X.M. Liu9, F. Zhang10, S.J. Romaniello11, A.D. Anbar11, Y. Peng12, Y. Cai13, J.D. Schiffbauer14, M. Meyer15, G.J. Gilleaudeau16, R.E. Plummer17, N.E. Sievers4,18, S. Goderis1, P. Claeys1
1Research Group of Analytical, Environmental and Geo- Chemistry, Vrije Universiteit Brussel, Brussels, Belgium. Email: Huan.Cui@vub.be, geohcui@gmail.com; ORCID ID: https://orcid.org/0000-0003-0705-3423, https://orcid.org/0000-0002-6666-7153, https://orcid.org/0000-0002-4585-7687
2State Key Laboratory of Palaeobiology and Stratigraphy, Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences, Nanjing, China
3Department of Geology and Earth System Science Interdisciplinary Center, University of Maryland, College Park, MD 20742, USA https://orcid.org/0000-0003-4129-6445
4Department of Geosciences, Virginia Tech, Blacksburg, VA 24061, USA. ORCID ID: https://orcid.org/0000-0003-4655-2663, https://orcid.org/0000-0001-8584-0428
5Trofimuk Institute of Petroleum Geology and Geophysics, Siberian Branch Russian Academy of Sciences, Novosibirsk 630090, Russia. ORCID ID: https://orcid.org/0000-0003-0797-1347, https://orcid.org/0000-0003-4875-5261, https://orcid.org/0000-0003-1880-5635
6Department of Geology and Geophysics, Novosibirsk State University, Novosibirsk 630090, Russia
7United States Geological Survey, Menlo Park, CA 94025, USA https://orcid.org/0000-0002-9770-6557
8División de Geociencias Aplicadas, IPICYT, San Luis Potosí 78216, Mexico. ORCID ID: http://orcid.org/0000-0003-4368-322X
9Department of Geological Sciences, University of North Carolina, Chapel Hill, NC 27599, USA. ORCID ID: https://orcid.org/0000-0001-7904-1056
10Natural History Museum of Denmark, University of Copenhagen, Copenhagen, Denmark. ORCID ID: https://orcid.org/0000-0003-3277-445X
11School of Earth and Space Exploration, Arizona State University, Tempe, AZ 85287, USA. ORCID ID: https://orcid.org/0000-0003-1582-6801, https://orcid.org/0000-0002-6015-7750
12Department of Geology and Geophysics, Louisiana State University, Baton Rouge, LA 70803, USA. ORCID ID: https://orcid.org/0000-0001-6458-5552
13State Key Laboratory of Continental Dynamics, Shaanxi Key Laboratory of Early Life and Environment, Department of Geology, Northwest University, Xi’an 710069, China. ORCID ID: https://orcid.org/0000-0002-9867-8156
14Department of Geological Sciences, University of Missouri, Columbia, Missouri 65211, USA. ORCID ID: https://orcid.org/0000-0003-4726-0355
15Department of Earth and Environmental Sciences, Harrisburg University, PA 17101, USA. ORCID ID: https://orcid.org/0000-0002-4164-5169
16Department of Atmospheric, Oceanic, and Earth Sciences, George Mason University, VA 22030, USA. ORCID ID: https://orcid.org/0000-0001-8131-8915
17Hydrology and Remote Sensing Laboratory, Beltsville Agricultural Research Center, US Department of Agriculture, Beltsville, MD 20705 USA. ORCID ID: https://orcid.org/0000-0002-6089-7791
18Nanoscale Characterization and Fabrication Laboratory, Virginia Tech, Blacksburg, Virginia 24061, USA.
| |
ABSTRACT
The terminal Ediacaran contains dramatic changes in biogeochemical cycles, many of which are closely coupled with evolutionary transitions in the corresponding fossil records. Dynamic redox conditions may have caused a profound impact on early animal evolution. Our work highlights the significance of integrated bio-, litho-, and chemo-stratigraphy in geobiology research of the deep time.
Keywords: Chemostratigraphy; Redox condition; Early macroorganisms; Biogeochemical cycles; Geobiology; Animal biomineralization; Alkalinity; Cloudina.
|
| |
RESUMEN
El Ediacárico terminal inlcuye una serie de cambios drásticos en los ciclos biogeoquímicos, muchos de los cuales se asocian con modificaciones evolutivas en los registros fósiles correspondientes. Entre ellos destacan las condiciones redox, las cuales pueden haber causado un impacto profundo en la evolución animal temprana. Este trabajo destaca el significado de la lito-, bio- y quimioestratigrafía en la investigación geobiológica del tiempo profundo.
Palabras clave: Quimioestratigrafía; Condiciones redox; Macroorganismos tempranos; Ciclos biogeoquímicos; Geobiología; Biomineralización animal; Alcalinidad; Cloudina.
|
IntroductionTOP
The terminal Ediacaran witnessed the first appearance of macroscopic organisms including the earliest biomineralizing animals in Earth history (Narbonne et al., 2012; Xiao et al., 2016). However, the biogeochemical context for this evolutionary milestone remains uncertain due to the absence of radiometric constraints for key events recorded in palaeontological and chemostratigraphic datasets, and to uncertainties in the correlation of these biogeochemical events worldwide. Moreover, soft-bodied Ediacara biotas are typically preserved as impressions in fine-grained siliciclastic rocks (Gehling, 1999), which offer limited opportunities for palaeoenvironmental analysis using geochemical tools. In contrast, two rare soft-bodied Ediacara assemblages are preserved in well-exposed marine carbonate successions of the Dengying Formation in South China (Chen et al., 2014) and the Khatyspyt Formation in Arctic Siberia (Grazhdankin et al., 2008), where comprehensive chemostratigraphic studies have recently been completed.
ResultsTOP
In the Gaojiashan Member of the Dengying Formation, integrated bio- and chemo-stratigraphic profiles 2019) show that the first appearance of the biomineralizing animal fossil Cloudina is closely associated with positive anomalies of δ13Ccarb, δ34Spyrite, and Sr/Ca values (Fig. 1). In contrast, S isotope values of carbonate-associated sulfate (δ34SCAS) remain steady throughout the succession, resulting in anomalously large (>70‰) sulfur isotope fractionations (δ 34SCAS-pyrite) in the lower half of the member that decline to ~30‰ in the upper half. This fractionation trend likely relates to changes in microbial communities, with sulfur disproportionation dominating in the lower interval, whereas microbial sulfate reduction was the principal metabolic pathway in the upper. We propose that the coupled palaeontological and biogeochemical transition may have coincided with an increase in terrestrial weathering fluxes of sulfate (evidenced by gypsum pseudomorphs), alkalinity (evidenced by Sr/Ca positive anomaly), and nutrients to the depositional basin, which stimulated primary productivity (evidenced by high δ13Ccarb values) and the spread of an oxygen minimum zone (evidenced by high δ15N and low δ238U values). Enhanced production and burial of organic matter is thus directly connected to the δ13Ccarb anomaly, and through bottom water anoxia likely promoting microbial sulfate reduction and pyritization as the main taphonomic pathway for Conotubus and other soft-bodied Ediacara biota. These results suggest that environmental and physiological pressures from an increase in seawater alkalinity - likely coupled with ecological pressures from predation (Hua et al., 2003, 2007) - set the stage for the evolutionary novelty of animal biomineralization.
|
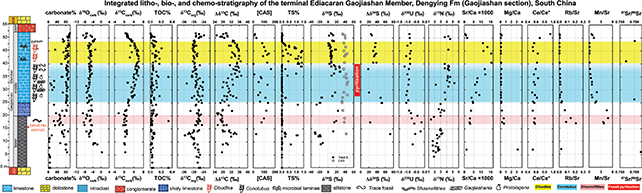 Figure 1.—‒Integrated litho-, bio-, and chemo-stratigraphy of the Gaojiashan Member, Dengying Formation at the Gaojiashan section, South China. Geochemical profiles include carbonate content (wt. %), carbonate oxygen isotopes (δ18Ocarb, ‰ VPDB), carbonate carbon isotopes (δ13Ccarb, ‰ VPDB), total organic carbon content (TOC), organic carbon isotopes (δ13Corg, ‰ VPDB), carbon isotope fractionations (Δδ13Ccarb-org), CAS concentration ([CAS] in ppm), total sulfur content (TS%, dominated by pyrite with trace amount of organic S), total sulfur isotopes in acidified residuals (δ34STS, ‰ VCDT), CAS sulfur (δ34SCAS, ‰ VCDT) isotopes, sulfur isotope fractionations (Δδ34SCAS-pyrite) assuming the δ34STS signals are dominantly derived from pyrite, uranium isotopes (δ238U, ‰ CRM145), nitrongen isotopes (δ15N, ‰ AIR), Sr/Ca, Mg/Ca, Ce anomaly (Ce/Ce*) calculated using the formula Ce/Ce* = CePAAS/([Pr]2PAAS / [Nd]PAAS), Rb/Sr, Mn/Sr, 87Sr/86Sr values measured from limestone beds or limestone nodules. BW = Beiwan, AD = Algal Dolomite. Data source: δ15N data , δ238U data , all the other data . Modified after Cui et al. (2016b, 2019). Figure 1.—‒Integrated litho-, bio-, and chemo-stratigraphy of the Gaojiashan Member, Dengying Formation at the Gaojiashan section, South China. Geochemical profiles include carbonate content (wt. %), carbonate oxygen isotopes (δ18Ocarb, ‰ VPDB), carbonate carbon isotopes (δ13Ccarb, ‰ VPDB), total organic carbon content (TOC), organic carbon isotopes (δ13Corg, ‰ VPDB), carbon isotope fractionations (Δδ13Ccarb-org), CAS concentration ([CAS] in ppm), total sulfur content (TS%, dominated by pyrite with trace amount of organic S), total sulfur isotopes in acidified residuals (δ34STS, ‰ VCDT), CAS sulfur (δ34SCAS, ‰ VCDT) isotopes, sulfur isotope fractionations (Δδ34SCAS-pyrite) assuming the δ34STS signals are dominantly derived from pyrite, uranium isotopes (δ238U, ‰ CRM145), nitrongen isotopes (δ15N, ‰ AIR), Sr/Ca, Mg/Ca, Ce anomaly (Ce/Ce*) calculated using the formula Ce/Ce* = CePAAS/([Pr]2PAAS / [Nd]PAAS), Rb/Sr, Mn/Sr, 87Sr/86Sr values measured from limestone beds or limestone nodules. BW = Beiwan, AD = Algal Dolomite. Data source: δ15N data , δ238U data , all the other data . Modified after Cui et al. (2016b, 2019).
|
|
In the Khatyspyt Formation, the profound sulfur isotope fractionation trend seen in the Dengying is preserved, with δ34Spyrite values rising from ca. - 20‰ to ca. 50‰ through an interval of general invariance in δ34SCAS (Fig. 2). The step function shift in sulfur isotope compositions is closely associated with the first occurrence of soft-bodied Ediacara biotas preserved in thinly bedded limestones, which suggests a possible link between seawater redox conditions and the distribution of these enigmatic macroscopic organisms. Based on multiple lines of sedimentological and geochemical evidence, we propose that the development of oceanic euxinia - which could be widespread in the continental margins due to enhanced oxidative weathering (delivering both nutrients and sulfate to ocean margins) in the terminal Ediacaran Period - may have locally prohibited the colonization of Ediacara-type organisms. The progressive secular transition from euxinic to non-euxinic and more habitable conditions may have allowed for the colonization of Ediacara-type and other macro-organisms in the upper part of the studied section of the Khatyspyt Formation.
|
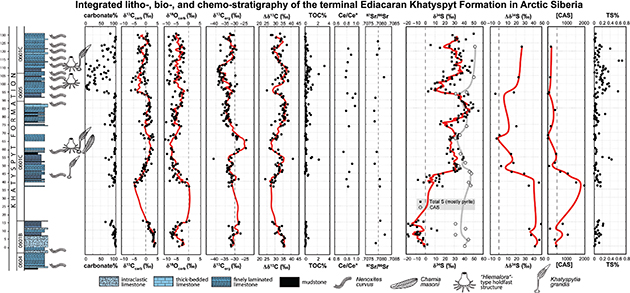 Figure 2.—‒Integrated litho-, bio-, and chemo-stratigraphy of the Khatyspyt Formation at the 0601 and 0605 sections, Arctic Siberia, Russia. Measured geochemical profiles include carbonate percentage (carbonate%), carbonate carbon (δ13Ccarb, ‰ VPDB) and oxygen isotopes (δ18Ocarb, ‰ VPDB), organic carbon isotopes (δ13Corg, ‰ VPDB), carbon isotope fractionations (Δδ13Ccarb-org), total organic carbon content (TOC, %), Ce anomaly (Ce/Ce*) calculated using the formula Ce/Ce* = [Ce]PAAS / ([Pr]2PAAS / [Nd]PAAS), strontium isotope ratios (87Sr/86Sr), total sulfur isotopes in acidified residuals (δ34STS, ‰ VCDT; including pyrite and trace amount of organic S), CAS sulfur isotopes (δ34SCAS, ‰ VCDT), sulfur isotope fractionations (Δδ34SCAS-pyrite) assuming the δ34STS signals are dominantly derived from pyrite, CAS concentration ([CAS] in ppm), and total sulfur content (TS%, dominated by pyrite with trace amount of organic S). Fitted lines represent five-point running average for δ13Ccarb and δ34STS data, and two-point running average for δ34SCAS, Δδ34S and [CAS] data. Modified after Cui et al. (2016a). Figure 2.—‒Integrated litho-, bio-, and chemo-stratigraphy of the Khatyspyt Formation at the 0601 and 0605 sections, Arctic Siberia, Russia. Measured geochemical profiles include carbonate percentage (carbonate%), carbonate carbon (δ13Ccarb, ‰ VPDB) and oxygen isotopes (δ18Ocarb, ‰ VPDB), organic carbon isotopes (δ13Corg, ‰ VPDB), carbon isotope fractionations (Δδ13Ccarb-org), total organic carbon content (TOC, %), Ce anomaly (Ce/Ce*) calculated using the formula Ce/Ce* = [Ce]PAAS / ([Pr]2PAAS / [Nd]PAAS), strontium isotope ratios (87Sr/86Sr), total sulfur isotopes in acidified residuals (δ34STS, ‰ VCDT; including pyrite and trace amount of organic S), CAS sulfur isotopes (δ34SCAS, ‰ VCDT), sulfur isotope fractionations (Δδ34SCAS-pyrite) assuming the δ34STS signals are dominantly derived from pyrite, CAS concentration ([CAS] in ppm), and total sulfur content (TS%, dominated by pyrite with trace amount of organic S). Fitted lines represent five-point running average for δ13Ccarb and δ34STS data, and two-point running average for δ34SCAS, Δδ34S and [CAS] data. Modified after Cui et al. (2016a).
|
|
ConclusionsTOP
In the Gaojiashan Member of the Dengying Formation, China, environmental and physiological pressures were linked to an increase in seawater alkalinity, probably coupled with ecological pressures from predation, setting the stage for the evolutionary novelty of animal biomineralization. In the Khatyspyt Formation of Arctic Siberia, the progressive secular transition from euxinic to non-euxinic and more habitable conditions may have allowed for the colonization of Ediacara-type and other macro-organisms.
ReferencesTOP
| ○ |
Chen, Z.; Zhou, C.; Xiao, S.; Wang, W.; Guan, C.; Hua, H. & Yuan X. (2014). New Ediacara fossils preserved in marine limestone and their ecological implications. Scientific Reports, 4: 4180. https://doi.org/10.1038/srep04180 |
| ○ |
Cui, H.; Grazhdankin, D.V.; Xiao, S.; Peek, S.; Rogov, V.I.; Bykova, N.V.; Sievers, N.E.; Liu, X.-M. & Kaufman, A.J. (2016a). Redox-dependent distribution of early macro-organisms: Evidence from the terminal Ediacaran Khatyspyt Formation in Arctic Siberia. Palaeogeography, Palaeoclimatology, Palaeoecology, 461: 122–139. https://doi.org/10.1016/j.palaeo.2016.08.015 |
| ○ |
Cui, H.; Kaufman, A.J.; Xiao, S.; Peek, S.; Cao, H.; Min, X.; Cai, Y.; Siegel, Z.; Liu, X.M.; Peng, Y.; Schiffbauer, J.D. & Martin, A.J. (2016b). Environmental context for the terminal Ediacaran biomineralization of animals. Geobiology, 14: 344–363. https://doi.org/10.1111/gbi.12178 |
| ○ |
Cui, H.; Xiao, S.; Cai, Y.; Peek, S.; Plummer, R.E. & Kaufman, A.J. (2019). Sedimentology and chemostratigraphy of the terminal Ediacaran Dengying Formation at the Gaojiashan section, South China. Geological Magazine, in press, https://doi.org/10.1017/S0016756819000293 |
| ○ |
Gamper, A. (2014). Global trends in nutrient dynamics during the Ediacaran-Cambrian period as revealed by nitrogen and carbon isotope trends. PhD Thesis, Freie University, Berlin, https://d-nb.info/1064307035/34 |
| ○ |
Gehling, J.G. (1999). Microbial mats in terminal Proterozoic siliciclastics: Ediacaran death masks. Palaios, 14: 40–57. https://doi.org/10.2307/3515360 |
| ○ |
Grazhdankin, D.V.; Balthasar, U.; Nagovitsin, K.E. & Kochnev, B.B. (2008). Carbonate-hosted Avalon-type fossils in arctic Siberia. Geology, 36: 803–806. https://doi.org/10.1130/G24946A.1 |
| ○ |
Hua, H.; Pratt, B.R. & Zhang L.Y. (2003). Borings in Cloudina shells: complex predator-prey dynamics in the terminal Neoproterozoic. Palaios, 18: 454–459. https://doi.org/10.1669/0883-1351(2003)018<0454:BICSCP>2.0.CO;2 |
| ○ |
Hua, H.; Chen, Z. & Yuan, X. (2007). The advent of mineralized skeletons in Neoproterozoic Metazoa - new fossil evidence from the Gaojiashan Fauna. Geological Journal, 42: 263–279. https://doi.org/10.1002/gj.1077 |
| ○ |
Narbonne, G.M.; Xiao, S.; Shields, G.A. & Gehling J.G. (2012). The Ediacaran Period. In: The Geologic Time Scale (Gradstein, F.M.; Ogg, J.G.; Schmitz, M.D. & Ogg, G.M. eds.). Elsevier, Boston, USA, 413–435. https://doi.org/10.1016/B978-0-444-59425-9.00018-4 |
| ○ |
Schiffbauer, J.D.; Xiao, S.; Cai, Y.; Wallace, A.F.; Hua, H.; Hunter, J.; Xu, H.; Peng, Y. & Kaufman, A.J. (2014). A unifying model for Neoproterozoic-Palaeozoic exceptional fossil preservation through pyritization and carbonaceous compression. Nature Communications, 5: 5754. https://doi.org/10.1038/ncomms6754 |
| ○ |
Xiao, S.; Narbonne, G.M.; Zhou, C.; Laflamme, M.; Grazhdankin, D.V.; Moczydłowska-Vidal, M. & Cui, H. (2016). Toward an Ediacaran time scale: Problems, protocols, and prospects. Episodes, 39: 540–555. https://doi.org/10.18814/epiiugs/2016/v39i4/103886 |
| ○ |
Zhang, F.; Xiao, S.; Kendall, B.; Romaniello, S.J.; Cui, H.; Meyer, M.; Gilleaudeau, G.J.; Kaufman, A.J. & Anbar, A.D. (2018). Extensive marine anoxia during the terminal Ediacaran Period. Science Advances 4, eaan8983. https://doi.org/10.1126/sciadv.aan8983 |