Short abstracts presented to the International Meeting on the Ediacaran System and the Ediacaran-Cambrian Transition (Guadalupe, Extremadura, Spain, October 17-24th, 2019)
Resúmenes cortos presentados en el Congreso Internacional sobre el Ediacárico y el tránsito Ediacárico-Cámbrico (Guadalupe, Extremadura, España, 17 al 24 de Octubre de 2019)
J.J. Álvaro1, S. Jensen2 (Eds.)
1Instituto de Geociencias (CSIC-UCM), Dr. Severo Ochoa 7, 28040 Madrid, Spain. Email: jj.alvaro@csic.es; ORCID ID: https://orcid.org/0000-0001-6294-1998
2Área de Paleontología, Facultad de Ciencias, Universidad de Extremadura, 06071 Badajoz, Spain. Email: soren@unex.es; ORCID ID: https://orcid.org/0000-0003-4573-9282
| |
PRESENTATION
The following document compiles the short contributions to the IMECT (International Meeting on the Ediacaran System and the Ediacaran-Cambrian Transition) meeting, celebrated in October 17-24th, 2019, in Guadalupe (Villuercas-Ibores-Jara Geopark). These abstracts have not been peer-reviewed and were only revised from a formal point of view by the Organizing Committee.
|
| |
PRESENTACIÓN
El siguiente documento recopila los resúmenes cortos presentados en el IMECT (International Meeting on the Ediacaran System and the Ediacaran-Cambrian Transition), celebrado del 17 al 24 de octubre de 2019 en Guadalupe (Geoparque Villuercas-Ibores-Jara). Estos resúmenes no han sido sometidos a revisión por pares y sólo fueron revisados por el Comité Organizador por lo que respecta a aspectos formales.
|
Diversification, behavioural traits and palaeoecology of burrows: a case study from the Cambrian Nagaur Sandstone, Marwar Supergroup, Rajashtan, IndiaTOP
S. Ahmad1, A. Srivastava2, M. Sharma1, S. K. Pandey1, A. H. Ansari1, P. Srivastava2
1Birbal Sahni Institute of Palaeosciences, 53 University Road, Lucknow, India. Email: shamim.ahmad@bsip.res.in
2Centre of Advanced Study, Department of Geology, University of Lucknow, India
The terminal Precambrian Era experienced numerous integrated changes in lithosphere-atmosphere-hydrosphere-biosphere, such as changes in crustal coverage, deep water oxygenation and the evolution of macroscopic biota. Deep water oxygenation and increased nutrient availability were key drivers in the development of Ediacara biota and acanthomorphic acritarchs. The distinct Ediacaran animal fossils disappeared before the onset of the Cambrian and paved the way for new more complex animals. Evolution of bioturbation activity is one of such development that has been recorded since latest Ediacaran times and had become more prominent in the Cambrian. The present study is focused on the significant aspects of the spatio-temporal variability in bioturbation features and its connection with the depositional environment.
To address this issue, six well-preserved ichnogenera have been identified and reviewed, which are Planolites, Palaeophycus, Bergaueria, Monocraterion, Skolithos and Treptichnus pedum. All these have been systematically studied from the Cambrian Nagaur Sandstone, Marwar Supergroup. Global occurrence and depositional niche of these trace fossils have been reviewed, to understand the corresponding palaeoecological changes over time and space. The ichnogenera Planolites and Skolithos and the ichnospecies Treptichnus pedum have been reviewed within the Cambrian Period, whereas Monocraterion, Berguaeria and Palaeophycus are reviewed throughout the whole Phanerozoic Era to know the consistency/inconsistency of dataset. Statistical analysis on the gathered dataset exhibits a trend between burrows and their occurrences in Cambrian and overlying Phanerozoic strata: i.e., the abundance of these burrows decreases in successive younger stratigraphic column of the Phanerozoic Era. This suggests the active population of burrow makers such as crustaceans, coelenterates, arthropods and biletarian organisms, directly proportional to the burrows in the early Cambrian, which was later progressively replaced by surface dwelling animals. Overall, their palaeoecology suggests that all six ichnogenera preserved in two different major depositional regimes, such as intertidal and subtidal environments. Thus, the burrowing activity better developed in intertidal than in subtidal regimes. The tendency of burrowing in the intertidal regime may be due to the presence of comparatively quite and stable water column with a better food web, whereas the subtidal regime is dominated by wave and storm activities, which either hinder the burrowing activity or effect the preservation. It must be noted that that primary purpose of burrowing was to get shelter from predator and hunting for food. Therefore, we propose any of these two constraints as becoming more moderate in the Furongian.
Unveiling the Cloudina-microbial reefs of a Cadomian retro-arc basin in West GondwanaTOP
J.J. Álvaro1, I. Cortijo2, S. Jensen3, M. Martí Mus3, T. Palacios3
1Instituto de Geociencias (CSIC-UCM), Dr. Severo Ochoa 7, 28040 Madrid, Spain. Email: jj.alvaro@csic.es
2Villuercas-Ibores-Jara UNESCO Global Geopark, Pintores 10, 10003 Cáceres, Spain. Email: icortijo@dip-caceres.es
3Área de Paleontología, Facultad de Ciencias, Universidad de Extremadura, 06071 Badajoz, Spain. Email: soren@unex.es, martimus@unex.es, medrano@unex.es
The Navalpino anticline of the Central Iberian Zone provides a unique case study of well-exposed late Ediacaran reefs. Drone images show their spatial arrangement as tens of cloudinid-microbial patch, biohermal and complex reefs, encased in a dominantly shaly succession. The latter was deposited on a perturbed block-platform placed in a Cadomian retro-arc basin fringing West Gondwana. Reefs display the traditional core/flank/inter-reef subdivision and developed in generally low-energy, clear-water substrates, between normal and storm wave base. Cores consist of millimetre-sized irregular mesoclots of dense microbial microsparite, locally interrupted by laminated (stromatolitic) fabrics, containing abundant Sinotubulites, Protolagena and Cloudina shells, the latter with upright preservation. Seafloor breakdown perturbations related to fissuring, hydrothermal influence and slope-apron breccia deposition led to nucleation and development of patch-reefs on unstable substrates. Their ecological succession is characterized by stabilization (pioneer)-to-colonization phases, the latter represented by Cloudina shells preserved upright and encased in microbial crusts, pointing to the active role of cloudinids as pioneer frame-building taxa.
The Precambrian-Cambrian transition in the PyreneesTOP
J.J. Álvaro1, J.M. Casas2, M. Padel3, S. Clausen4
1Instituto de Geociencias (CSIC-Universidad Complutense de Madrid), Dr. Severo Ochoa 7, 28040 Madrid, Spain. Email: jj.alvaro@csic.es
2Dept. de Dinàmica de la Terra i de l’Oceà - Institut de Recerca Geomodels, Universitat de Barcelona, Martí i Franquès s/n, 08028 Barcelona, Spain. Email: casas@ub.edu
3Bureau de Recherches Géologiques et Minières (BRGM), 3 Avenue Claude Guillemin, 45100 Orléans, France. Email: m.padel@brgm.fr
4Unité Mixte de Recherche (UMR), Université de Lille, 8198 EEPCNRS, Université de Lille 1, Bâtiment SN5, Avenue Paul Langevin, 59655 Villeneuve d’Ascq Cedex, France. Email: sebastien.clausen@univ-lille1.fr
The Pyrenees is an Alpine intracontinental fold and thrust belt that resulted from the convergence between the Iberian and European plates from Late Cretaceous to Oligocene times. In the Pyrenees, rocks ranging in age from late Neoproterozoic to Mississippian form an elongated strip in the backbone of the chain, unconformably overlain by Mesozoic and Cenozoic rocks. This pre-Variscan basement lies geographically disconnected from neighbouring outcrops that also belong to the eastern branch of the Ibero-Armorican Arc, such as the Catalan Coastal Range to the South, the Mouthoumet and Montagne Noire (southern French Massif Central) to the North, and Sardinia to the East.
The lower part of this pre-Variscan succession is made up by a pre-Upper Ordovician succession, more than 3000 m thick, which crops out extensively in the central and eastern Pyrenees and contains the Ediacaran-Cambrian transition. However, precise location of this boundary has been hampered by the monotonous character of an important part of the succession, the lack of a well-preserved fossiliferous record and the complex macrostructure. The scarcity of biostratigraphic data in the Ediacaran-Lower Ordovician succession has been partly compensated by reporting of geochronological U/Pb dating of zircon of interbedded metavolcanic rocks. This, together with detailed field work, allow to propose recently a new stratigraphic framework for this lower part of the pre-Variscan sequence, from bottom to top: The Canaveilles Group, with the Nyer, Olette and Pic de la Clape formations and the Jujols Group, with the Err, Valcebollère and Serdinya formations. An Ediacaran-early Cambrian age can be proposed for the Canaveilles Group and a broad Cambrian-earliest Ordovician age for the entire Jujols Group. The Ediacaran-Cambrian transition should be located within the Pic de la Clape Formation, which includes a succession of genetically related volcanic pulsations linked to episodes of carbonate production. Geochronological data also constrain the depositional age of the Ediacaran succession in the Eastern Pyrenees, from ca. 570 to 542 Ma, although the oldest ages obtained from metavolcanic rocks should be regarded as a minimum because of a thick series cropping out below these rocks.
Kimberichnus: Is Kimberella really a trace maker?TOP
J.B. Antcliffe1
1Université de Lausanne, Institut des Sciences de la Terre, Bâtiment Géopolis, UNILMouline, CH-1015 Lausanne, Switzerland. Email: jonathan.antcliffe@unil.ch
The taphonomy of Ediacaran age soft bodied fossils has been extensively studied because global soft fossil preservation is highly unusual at all other times in Earth history. Ediacaran fossils are typically the results of impressions made by soft and flexible organisms in siliciclastic sediments. This mode of preservation is almost entirely absent from the fossil record from the end of the Cambrian Period onwards. Kimberella is known from rocks from the Flinders Ranges, South Australia and the White Sea region of Russia where the fossils are formed by upper bed lithification, which forms negative fossil moulds on the underside of the overlying bed. Consequently, the top surface of the organism is preserved as the impression (negative hyporelief). Microbial mats are central to the preservation model as microbial mats covered the seafloor in the Ediacaran period.
The microbial mats aid in rapid and early lithification (e.g. through trapping a binding), stabilise soft tissue, mediate pyritisation and diagenesis, and may even be moulding the organism in life. Fossil bedding surfaces covered in micro-bial mats have a wrinkled texture with a range of characters that are distinctive such as pustules, over-steepened ridges, contortion, tea-ring and folding of the microbialite, suspended sediments grains, and microscale concentrations of mine-rals such as mica flakes in the upper layers and pyrite grains beneath. These microbial mats are pervasive on bedding planes at the Flinders Ranges and White Sea fossil sites. The range of their textures and structures however is poorly documented and this new analysis shows that there is a spectrum of structures that grade in to each other, ranging from densely packed bundles of fibres with large tangled pustules to sparsely arranged filamentous structures as the mat first colonises a bedding surface. In the intergradation between these two extremes are a huge range of complex structures formed as the filaments start to overlap and interact, of which the most common are cross hatching fibrous structures as the microbial mat begins to knit toge-ther. The Kimberichnus “trace fossil” falls into this continuum closer to the surface colonisation end of the spectrum, showing cross-hatching, filament bifurcation, overlapping of filaments, and filament tangling which would be impossible in a trace fossil (Fig. 1). The Kimberichnus trace is best described as a variant of a well-known microbialite, and does not have any particular association with Kimberella body fossils. It is commonly found alone on bedding planes, or intergrading into other microbialite textures, or even alongside other Ediacaran body fossils. Kimberella does not appear to have been actively motile and it does not seem to have been leaving feeding traces, as a result, the iconic interpretation of this fossil as a microbial mat eating mollusc should be considered doubtful.
|
 Figure 1.—(overleaf) A. SAM P35655 Kimberichnus from the Flinders Ranges, South Australia showing slight cross-hatching of filaments, tangling of filaments, and many bifurcations of filaments on a sparsely occupied bedding plane. B. SAM P35663 Kimbe-richnus from the Flinders Ranges, South Australia showing extensive cross-hatching of filaments on a bedding plane with abundant and densely packed microbialites. Scale bar divisions are 1 mm. Figure 1.—(overleaf) A. SAM P35655 Kimberichnus from the Flinders Ranges, South Australia showing slight cross-hatching of filaments, tangling of filaments, and many bifurcations of filaments on a sparsely occupied bedding plane. B. SAM P35663 Kimbe-richnus from the Flinders Ranges, South Australia showing extensive cross-hatching of filaments on a bedding plane with abundant and densely packed microbialites. Scale bar divisions are 1 mm.
|
|
Sediment rheology explains the Ediacara biota preservationTOP
I. Bobrovskiy1, A. Krasnova2,3, J.J. Brocks1
1Australian National University, Australia. Email: ilya.bobrovskiy@anu.edu.au
2Borissiak Paleontological Institute, Russian Academy of Sciences, Russia.
3Lomonosov Moscow State University, Russia.
The Ediacara biota (571-539 Ma) represents the oldest complex large organisms in the fossil record, providing a bridge between largely microbial ecosystems of the Precambrian and the animal dominated world of the Phanerozoic. However, the nature of most Ediacaran organisms remains unclear, not least due to their enigmatic non-actualistic preservation. We show that Flinders-style fossilization of Ediacaran organisms was promoted by unusually prolonged conservation of organic matter, coupled with differences in rheological behaviour of the over- and underlying sediments. In contrast to accepted models, cementation of overlying sand was not critical for fossil preservation, which is supported by the absence of cement in unweathered, unmetamorphosed White Sea specimens and observations of soft sediment deformation in both White Sea and South Australian specimens. This model has been confirmed by laboratory simulations, which produced Ediacaran-like negative hyporelief impressions at the base of sand layers. The rheological model implies that Ediacaran fossils do not necessarily reflect the external shape of the organism. In many cases, the impressions repeat the morphology of a soft external or internal organic ‘skeleton’. This mechanism provides new constraints on biological interpretations of the Ediacara biota.
Regional nutrient decrease drove redox stabilisation and metazoan diversification in the late Ediacaran Nama Group, NamibiaTOP
F.T. Bowyer1+, A.J. Shore1*, R.A. Wood1, L.J. Alcott2, A.L. Thomas1, I.B. Butler1, A. Curtis1, S. Hainanan3, S. Curtis-Walcott4, A.M. Penny5, S.W. Poulton2
1University of Edinburgh, School of GeoSciences, James Hutton Road, Edinburgh, EH9 3FE, UK.
2University of Leeds, School of Earth and Environment, Leeds, LS2 9JT, UK.
3Ministry of Mines and Energy, 6 Aviation Road, Private Bag 13297, Windhoek, Namibia.
4Department of Physics, Imperial College, London SW7 2AZ.
5Finnish Museum of Natural History, University of Helsinki, Jyrängöntie 2, 00560 Helsinki, Finland.
+Current address: University of Leeds, School of Earth and Environment, Leeds, LS2 9JT, UK
The late Ediacaran witnessed an increase in metazoan diversity and ecological complexity, marking the inception of the Cambrian Explosion. To constrain the drivers of this diversification, we combine local redox and nutrient data for two shelf transects, with a complete inventory of biotic diversity and distribution from the Nama Group, Namibia (~550 to ~538 Million years ago; Ma). Unstable marine redox conditions characterise all water depths in inner to outer ramp from ~550 to 547 Ma when the first skeletal metazoans appeared, but a marked deepening of the redoxcline and a reduced frequency of anoxic incursions onto the inner to mid-ramp is recorded from ~547 Ma onwards, with full ventilation of the outer ramp by ~542 Ma. Phosphorus speciation data show that initial anoxic ferruginous conditions promoted the drawdown of bioavailable P, with productivity constrained by limited P recycling back to the water column. A long-term decrease in nutrient delivery from continental weathering, coupled with a possible decrease in upwelling, led to the gradual ventilation of the Nama Group basins. This in turn decreased anoxic recycling of bioavailable phosphorus to the water column, promoting the development of stable oxic conditions and the radiation of new mobile taxa.
Sediment bulldozing and the Ediacaran-Cambrian transition: Evidence from the Nama Group of NamibiaTOP
L.A. Buatois1, J. Almond2, M.G. Mángano1, S. Jensen3, G.J.B. Germs4
1Department of Geological Sciences, University of Saskatchewan, 114 Science Place, Saskatoon, Saskatchewan, S7N 5E2, Canada. Email: luis.buatois@usask.ca.
2Natura Viva cc, Cape Town 8001, South Africa.
3Área de Paleontología, Facultad de Ciencias, Universidad de Extremadura, E-06006, Badajoz, Spain.
4Department of Geology, University of the Free State, Bloemfontein, South Africa.
Ediacaran benthic marine ecosystems were dominated by extensive microbial mats that effectively sealed the sediment from the water column. Ediacaran ichnofaunas are characterized by non-penetrative, non-specialized horizontal trails interpreted as produced by vagile bilaterian metazoans that exploited organic matter concentrated within microbial mats. However, there is growing evidence that this situation started to change in the terminal Ediacaran with the appearance of other style of animal-substrate interactions. The Ediacaran-Fortunian Nama Group of Namibia, with its thick, laterally continuous, virtually undeformed and relatively well-dated shallow-marine successions, is proving to be a key unit to delineate these evolutionary innovations. In particular, the recently introduced ichnotaxon Parapsammichnites pretzeliformis, which occurs in terminal Ediacaran strata of the Spitskop Member of the Urusis Formation, provides uncontroversial evidence of sediment bulldozing. This ichnotaxon consists of unilobate to bilobate horizontal to subhorizontal trace fossils forming scribbles, circles and, more rarely, open spirals and meanders, and displaying an internal structure indicative of active fill. The occurrence of this ichnospecies in terminal Ediacaran strata of the Nama Group provides the earliest clear representatives so far recorded of sediment bulldozing. This activity may have had a negative impact on suspension-feeding and/or osmotroph communities, as well as on matgrounds, representing early examples of ecosystem engineering and trophic-group amensalism. Efficient burrowing may indicate coelom development most likely linked to metazoan body-size increase. The occurrence of sediment bulldozers may have promoted the establishment of gradients in horizontal and vertical distribution of organic material in connection with spatially heterogeneous environments on the sea floor at a critical time in Earth evolution.
The evolutionary timing of the origin of the animalsTOP
G.E. Budd1
1Uppsala University, Dept of Earth Sciences, Palaeobiology, Uppsala, Sweden. Email: Graham.budd@pal.uu.se
When the animals arose and diversified continues to be controversial. Here I critically review the stratigraphic and other evidence in the Ediacaran to Cambrian interval to attempt to establish a robust timescale for this critical evolutionary time.
The evidence for a deep (i.e. pre-Ediacaran) origin of the crown group animals continues to weaken, and birth-modelling animals support this contention. As a large clade, animals are likely to have diversified very quickly, and there can be little support for a “phylogenetic fuse” type model for their origins. The evidence of the fossil record is in particular incompatible with a deep bilaterian origin, and although bilaterians may well have begun to radiate just before the end of the Ediacaran, their major diversification is undoubtedly an event of the early Cambrian. Critical time points of the Cambrian itself, including the age of the base, the age of the first trilobites, and the age of the early exceptionally-preserved biota, all remain relatively unconstrained when the evidence is critically examined. Despite this fog of uncertainty, the overall timing of the animal radiation seems relatively clear, and it is thus possible to examine its major features against a backdrop of the evolving planet, not least the complex global tectonics of the Ediacaran-Cambrian. Such a reconstruction is not completely compatible with most molecular clock estimates of animal origins even when most generously interpreted, and possible reasons for this need to be investigated further before agreement can be reached on this most significant of evolutionary events.
Bagworld: constructional and functional anatomy of Ediacaran rangeomorphsTOP
N.J. Butterfield1
1Department of Earth Sciences, University of Cambridge, Cambridge, UK CB2 3EQ. Email: njb1005@cam.ac.uk
Ediacaran rangeomorphs are the first substantially macroscopic organisms to appear in the fossil record, but their underlying biology remains problematic. Although demonstrably heterotrophic, their current interpretation as osmotrophic consumers of dissolved organic carbon (DOC) is incompatible with the inertial (> Re) and advective (> Pe) fluid-dynamics accompanying macroscopic length-scales. The key to resolving rangeomorph feeding and physiology lies in their underlying construction. Taphonomic analysis of three-dimensionally preserved Charnia from the White Sea identifies the presence of large, originally water-filled compartments that served both as a hydrostatic exoskeleton and semi-isolated digestion chambers capable of processing recalcitrant substrates – almost certainly in conjunction with a resident microbiome. At the same time, the hydrodynamically exposed outer surface of macroscopic rangeomorphs would have dramatically enhanced both gas exchange and food delivery. A bag-like epithelium filled with transiently circulated seawater offers an exceptionally efficient means of constructing a multicellular DOC-consuming organism. Such a bodyplan is broadly comparable to that of anthozoan cnidarians, minus their clearly derived predatory adaptations such muscle, cnidae, tentacles and a centralized mouth. Along with a variety of other essentially bag-like fossils of Ediacaran and early Palaeozoic age, rangeomorphs can be reliably interpreted as total-group eumetazoans, most likely colonial, pre-predatory, stem-group cnidarians.
Biostratigraphy and chemostratigraphy of the Ediacaran-Cambrian transition in Sonora, Mexico: review and perspectivesTOP
S. Clausen1, L. Devaere1, J.P. Sosa León2, J.J. Palafox Reyes2, B.E. Buitrón-Sánchez3, D. Vachard4
1Université de Lille I, CNRS, UMR 8198 - Evo-Eco-Paleo, F-59000 Lille, France. Email: Sebastien.Clausen@univ-lille1.fr, lea_devaere@hotmail.fr
2Universidad de Sonora, Departamento de Geología, Boulevard Luis Encinas y Rosales, 83000 Hermosillo, Sonora, Mexico.
3Universidad Nacional Autónoma de México, Instituto de Geología, Departamento de Paleontología, Ciudad Universitaria, Delegación Coyoacán, 14510 México D.F., Mexico.
41 Les Tilleuls, 59152 Gruson, France.
The palaeontological record of the Ediacaran-Cambrian (E-C) transition, particularly of the Terreneuvian, remains poorly documented in Laurentia. Ediacaran fossils are relatively rare in the Great Basin despite extensive exposures of Proterozoic strata. In addition, except in the Northern Territories of Canada and possibly in the Great Basin (California and Nevada), all described first appearance data of Cambrian skeletonized faunas are post-trilobitic. As a consequence, no detailed biostratigraphic subdivisions have been proposed for the “pre-trilobitic” interval of Laurentia, and constraining the E-C boundary interval has remained difficult in most areas of this major palaeocontinent. Beside stratigraphic difficulties, such uncertainties impact our understanding of the triggers of this critical interval in Earth History. Proposed geodynamic triggers include the break up of Rodinia, generally estimated to end at about 555-550 Ma, but which rift-associated transform-faults might have remained episodically active till 525 Ma in the southwestern palaeomargin of Laurentia. In this context, the E-C succession in northwestern Sonora, which was located on the western margin of Laurentia, is unique. It records a thick, significant volcano-sedimentary episode interrupting a thick mixed carbonate-siliciclastic succession, and attesting for a still intense geodynamic activity in this area. Nevertheless, the exact duration and context of this episode remain unknown, as a result of which, the chronostratigraphy of the E-C succession remains unresolved despite abundant studies. Indeed, Ediacaran tubular fossils (possibly including the youngest cloudinids worldwide), Treptichnus pedum ichnofossils, Cambrian small shelly fossils, and later archaeocyaths and trilobites have all been independently described from Sonora. More recently, a δ13C chemostratigraphic characterisation of the E-C boundary interval was also proposed. However, the exact and relative positions of these proxies remain questionable as (i) always independently referred to an inherited schematic and over-synthetic column, and (ii) using an unstable lithostratigraphic scheme. To contribute overcoming these issues, a new study has been undertaken recently, including detailed chemostratigraphic, biostratigraphic and sedimentological analysis of different but key sections in Caborca area (Sonora). The first results demonstrate most regional stratigraphic schemes need to be emended. A significant and short-lived negative δ13C anomaly is recorded in all studied sections, in the same exact position upward revised cloudinid occurrences, and may mark the E-C boundary, allowing its correlation in the Great Basin. First recorded SSF assemblages demonstrate that most of the volcano-sedimentary interval, which geodynamic origin are still problematic, may not be Ediacaran to lowest Cambrian as previously stated, but instead Cambrian Stage 2 to 3, arguing for a major E-C regional discontinuity. The delayed first occurrence of Treptichnus pedum, reported from this interval, should not be used regionally as a marker of the E-C boundary.
Conodonts of the Deadwood Formation (Furongian), Saskatchewan, Canada and their biostratigraphic potentialTOP
W.J.E. Crabbe1, M.A. Purnell1, T.H.P. Harvey1
1Centre for Palaeobiology Research, School of Geography, Geology and the Environment, University of Leicester, Leicester, Leicestershire, LE1 7RH, UK. Email: wc117@leicester.ac.uk
Furongian biostratigraphy is typically dominated by trilobite biozonation schemes. However, the middle to upper Cambrian of subsurface Saskatchewan, Canada, is devoid of a diagnostic shelly macrofauna, except for a brachiopod assemblage from southwestern Saskatchewan (Robson et al., 2003). This makes a defined biostratigraphical framework problematic. Here we present a large and diverse assemblage of flattened, carbonaceous paraconodonts recovered from drillcore of the Deadwood Formation from near Regina, Saskatchewan (Butterfield & Harvey, 2012; see Fig 2).
|
 Figure 2.—Carbonaceous paraconodonts from the upper Cambrian Deadwood Formation of Saskatchewan, Canada. Scale bar = 100 µm. Figure 2.—Carbonaceous paraconodonts from the upper Cambrian Deadwood Formation of Saskatchewan, Canada. Scale bar = 100 µm.
|
|
The Deadwood paraconodont assemblage includes the first occurrences of Serratocambria and Proacodus outside of Swedish assemblages (see Müller & Hinz, 1991) as well as Westergaardodina concamerata and W. curvata which occur in the Furongian of Sweden, and W. tricuspidata which has been recovered from the Furongian of Antarctica, Poland, South China, South Korea and Sweden. Two apparently new Westergaardodina morphotypes have also been recovered. Additionally, a large and diverse assemblage of coniform paraconodont taxa is recorded, including Furnishina, Problematoconites, Prooeneotodus, Muellerodus and Trolmenia. This is the most diverse and well preserved paraconodont fauna recovered from Canada and therefore provides the opportunity for comparison with other conodont assemblages around the world. The overall signal from the Deadwood paraconodonts is broadly equivalent to conodont occurrences within the Protopeltura and Peltura superzones in Sweden, suggesting a position within the upper Furongian Series for the Deadwood assemblage. This study has highlighted the potential of paraconodonts for providing biostratigraphic resolution in strata that lack typically diagnostic taxa. This is the first time a paraconodont biozonation has been applied to siliciclastic strata and further analysis of siliciclastic Cambrian successions may yield further assemblages that could help to provide a more robust stratigraphical framework for the Furongian.
The early fossil record of Euarthropoda and the Cambrian ExplosionTOP
A.C. Daley1, J.B. Antcliffe1, H.B. Drage2, S.Pates2
1Institute of Earth Sciences, University of Lausanne, Géopolis, CH-1015 Lausanne. Email: allison.daley@unil.ch
2Department of Zoology, University of Oxford, The Tinbergen Building, South Parks Road, Oxford, OX1 3PS, UK.
As the most abundant and diverse animal phylum, eurarthropods have been major components of animal ecosystems for over 500 million years. Euarthropod fossils have also been key for examining the dynamics of the rapid early radiation of animals during the Cambrian Explosion. This event is documented by the Cambrian fossil record, however Precambrian ancestors have long been sought. Here we use the early fossil record of euarthropods as a model to explore the quality of fossil data as it relates to the Cambrian Explosion. Numerous types of fossil preservation, including soft-bodied macrofossils from Burgess Shale-Type (BST) localities, biomineralised exoskeletons, microfossils, and trace fossils are compared and contrasted across the Ediacaran-Cambrian boundary to constrain when euarthropods first evolved.
BSTs provide the most complete metazoan example of phylum-level anatomical construction in the euarthropod stem lineage during the Cambrian from 518 million years ago (Ma). The stem lineage includes non-biomineralized groups such as Radiodonta (e.g. Anomalocaris) that provide insight into the step-by-step construction of euarthropod morphology, including the exoskeleton, biramous limbs, segmentation, and cephalic structures. Trilobites are crown group euarthropods that appear in the fossil record at 521 Ma, before the stem lineage fossils, implying a ghost lineage that needs to be constrained. These constraints come from the trace fossil record, which show the first evidence for total group Euarthropoda (Cruziana, Rusophycus, etc.) at around 537 Ma. A deep Precambrian root to the euarthropod evolutionary lineage is disproven by a comparison of Ediacaran and Cambrian lagerstätten. BSTs from the latest Ediacaran Period (e.g. Miaohe Biota, 550 Ma) are abundantly fossiliferous with algae but completely lack animals, which are also missing from other Ediacaran windows, such as phosphate deposits (e.g. Doushantuo, 560 Ma). Furthermore, euarthropod candidate fossils from the Ediacaran Biota, such as Spriggina and Parvancorina, cannot be substantiated because they lack definitive euarthropod anatomical features. This constrains the appearance of the euarthropod stem lineage to no older than 550 Ma. While each of the major types of fossil evidence (BSTs, trace fossils, and biomineralised preservation) have their limitations and are incomplete in different ways, when taken together they allow a coherent picture to emerge. In congruence with analyses from molecular paleobiology, our comprehensive fossil data set suggests an entirely Cambrian origin and subsequent radiation for total group Euarthropoda.
Ichnotaxobases of trace fossils from the Corumbá Group - Tamengo and Gauicurus formations, BrazilTOP
C.C.Q. Diniz1, J.M. Leme2, R. Gandini3
1Programa de Pós-Graduação em Geociências (Geoquímica e Geotectônica), Instituto de Geociências, Universidade de São Paulo, Brazil. Email: cleber.diniz@hotmail.com.br
2Instituto de Geociências, Departamento de Geologia Sedimentar e Ambiental, Universidade de São Paulo, Brazil.
3Pós-doctoral reserarcher, Instituto de Geociências, Programa de Pós-Graduação em Geociências (Geoquímica e Geotectônica), Universidade de São Paulo, Brazil.
The presence of ichnofossils in the Ediacaran is already consolidated with the records especially of horizontal trails, but with low ichnodiversity and ichnodisparity with simpler morphology. The Ediacaran ichnofossils occupied both shallower marine environments in offshore-transition and offshore areas where their abundance and diversity reach the maximum, and to a lesser extent deep-marine environments in shelf areas. Ichnotaxobases are significant and variable morphological features of a trace that are directly connected to their behaviors. There are five main Ichnotaxobases: General form, Wall and lining, Fill, branching and spreiten. The Corumbá Group it is localized in Mato Grosso do Sul State in Brazil, surfaces in the southern portion of the Paraguay Fold Belt, in which the Guaicurus Formation corresponds to its top followed by the Tamengo, Bocaina, Cerradinho and Cadiueus formations. The Tamengo Formation age 542.37 ± 0.32 Ma have carbonaceous facies intercalated with packages of pelites, recorded with metazoan macrofossils, such as Corumbella werneri, Cloudina lucianoi, Paraconularia sp., vendotaenids, macroalgae and trace fossils. The Guaicurus Formation is composed by pelitic terrigenous sedimentation characterized by easy weathering of shales, recorded with algae fossils named as Eoholynia corumbensis and ichnofossils. It is fundamental to analyze the Ichnotaxobases of the trace fossils founded in Brazil, making it able to propose their behavior in specific environmental factors. Commonly the traces are horizontal to sub-horizontal burrows having a boundary by dust film or unlined, with active fill or massive passive fill but without branching and spreiten, with depauperate individuals, low diversity and encounter isolated, associated with Pascichnia ethology reflecting behaviors linked to feeding and locomotion altogether of detritus feeding in the surface such as grazing strategies or Fodinichnia ethology in which contemplates behaviors related to feeding and dwelling of deposit feeders that developed underground mining strategies. But in some cases further analysis are still important because only based in Ichnotaxobases can lead to mistaken interpretations, been able to be body fossil of tubular organisms or not biogenic at all, CT-scan analysis can help to answer questions that differentiate a body fossil from a trace fossil, because can show their full extension, the width along the structure, or the bioturbation within the lamination of the rock. The samples here defined as Pascichnia and Fodinichnia, fit in an environment of low energy, the presence of horizontal traces slightly entering the sediment in the seafloor can be attributed for the lack of infaunalization on Ediacaran, but also because of environmental factors, all the needs for the organism was in the surface, with high food supply from the microbial mats, but low oxygen rate that could only covers the surface of the seafloor, the organism could not cross to deeper areas. These factors can be attributed even to offshore zones as for shelf zones. The sedimentary structures of Guaicurus Formation with fined granulometry, millimetric ripples lamination that faded away to form plan-parallel structures and subtle bed forms imply calm marine environment deposited by suspension fall out. Therefore, both the characteristics presented by the ichnofossils and the sedimentary structures of the Corumbá Group, imply a deep marine environment closely to storm wave base such as lower offshore to shelf, represented by traces more isolated and superficial to the seafloor, and sporadic when was arise in food supply and / or oxygenation brought by the influence of currents, increases the number of grazing feeding and the depth at which producers can achieve.
Funding Projects: FAPESP (2016/ 06114-6); (2017/25433-8).
Developmental palaeobiology and phylogenetic affinities of the Ediacaran rangeomorphsTOP
F.S. Dunn1,2,3, C.G. Kenchington4, D.G. Grazhdankin5,6, P.R. Wilby3, P.C.J. Donoghue2, A.G. Liu4
1Oxford University Museum of Natural History, University of Oxford, Parks Road, Oxford, OX1 3PW, UK. Email: frances.dunn@oum.ox.ac.uk
2School of Earth Sciences, University of Bristol, Life Sciences Building, 24 Tyndall Avenue, Bristol, BS8 1TQ, UK.
3British Geological Survey, Nicker Hill, Keyworth, Nottingham, NG12 5GG, UK.
4Department of Earth Sciences, University of Cambridge, Downing Street, Cambridge, CB2 3EQ, UK.
5Trofimuk Institute of Petroleum Geology and Geophysics, Prospekt Akademika Koptyuga 3, Novosibirsk, 640090, Russia.
6Novosibirsk State University, Pirogova Street 1, Novosibirsk, 630090, Russia.
Ediacaran rangeomorphs possess a unique branching architecture and are amongst the most spatially and temporally widespread members of the Ediacaran macrobiota. Despite being plausibly interpreted as total-group metazoans, many aspects of their biology, as well as their unique phylogenetic placement, remain elusive. Historically, this has meant that rangeomorphs have not been considered in attempts to rationalise between different patterns of character acquisition in early-diverging metazoan clades. We reassess the anatomy, development and phylogenetic affinities of Rangeomorpha by examining a subset of taxa sampling the range of diversity within this group.
We report the developmental patterns of three rangeomorphs; Charnia masoni (uniterminal), Fractofusus (biterminal) and Hylaecullulus fordii (multiterminal), which sample the morphological variation of this group. These data enable us to conclude that the axial arrangements of rangeomorphs are both complex and disparate. A new homology scheme based upon our findings suggests that the classical uniterminal, biterminal and multiterminal groupings of taxa within the Rangeomorpha may not be founded in biological reality.
Finally, we present the results of a phylogenetic analysis concerning the affinities of Rangeomorpha and conclude they are crown-group animals, belonging to the stem-group Eumetazoa. These new data allow us to make inference about the evolution of eumetazoan character states and extends the fossil record of the Eumetazoa beyond 571 million years ago.
Geochemical and Nd-isotope features of Iberian sedimentary series. A key to interpret the Ediacaran-Cambrian peri-Gondwanan tectonic settingTOP
J.M. Fuenlabrada1, R. Arenas2, R. Díez Fernández3, S. Sánchez Martínez2, E. Rojo Pérez2, A.P. Pieren3
1Unidad de Geocronología (CAI de Ciencias de la Tierra y Arqueometría), Universidad Complutense, 28040 Madrid, Spain. Email: jmfuenla@ucm.es
2Departamento de Mineralogía y Petrología e Instituto de Geociencias (UCM, CSIC), Universidad Complutense, 28040 Madrid, Spain. Email: rarenas@ucm.es, s.sanchez@geo.ucm.es, e.rojo@ucm.es
3Departamento de Geodinámica, Estratigrafía y Paleontología, Universidad Complutense, 28040 Madrid, Spain. Email: rudiez@ucm.es, apieren@ucm.es
The Iberian Massif presents an extensive record of Ediacaran-Cambrian sedimentary rocks deposited along the African margin of Gondwana. They were involved in the Variscan Orogen, where they appear in different units both in autochthonous domains and included in large allochthonous complexes. These successions were originally formed in several E-W locations along the margin of Gondwana, and were later approached during dextral Variscan convergence. Consequently, the Iberian sedimentary successions bring an excellent opportunity to investigate the tectonic setting and Nd isotopic sources of the Ediacaran-Cambrian transition, and also the original palaeogeographic position of the Iberian terranes along the palaeomargin of Gondwana. In the NW of the Iberian Massif, the Basal Allochthonous Units of the Malpica-Tui Complex have been interpreted as an external section of the margin of Gondwana. The Basal Units represent a terrane with continental affinity composed of two metasedimentary sequences, different in age and composition. The lower metasedimentary sequence consists of a thick pile of metagreywackes deposited in the latest Neoproterozoic, while the upper succession is formed mainly by mica schists with a Furongian depositional age. Located under the allochthonous complexes, the Central Iberian Zone (CIZ) represents an autochthonous terrane interpreted as an inner section of the Gondwanan margin. CIZ presents a wide range of pre-Ordovician shales and sandstones distributed in two units: (i) the Lower Unit is mainly composed of monotonous successions of sandstones and shales, whose sedimentation took place over the Neoproterozoic; while (ii) the Upper Unit presents a greater lithological variety with a dominantly pelitic character, whose sedimentary facies suggest a more complex evolution of the sedimentary basins. Within the Upper Unit, the Pusa Group is mainly composed of shales deposited in early Cambrian times.
The Ediacaran greywacke successions from the Basal Allochthonous Units and the Central Iberian Zone share geochemical features typical for an active margin setting as the most probable depositional environment. Discrimination diagrams for the tectonic setting using trace elements point to a sedimentation influenced by an evolved volcanic arc, probably built over an extended and thinned continental basement. By contrast, the Cambrian siliciclastic successions show a greater recycled character and present similar compositions to those observed in common passive margins. Their highly homogeneous geochemical patterns confirm their recycled character and the greater influence of the sedimentary processes over the final chemical composition. These geochemical features suggest the development of the Ediacaran sedimentary series as related to the opening and early evolution of a peri-Gondwanan back-arc basin. The geochemical information provided for the siliciclastic rocks confirms the relatively closeness of the sedimentary basins to source areas, which is also consistent with the immaturity observed in the Ediacaran rocks. The widening of this back-arc basin over Cambrian times, led to a greater contribution from nearby cratonic areas. This stage in the evolution of the peri-Gondwanan realm also coincides with a decrease of the magmatic activity in the volcanic arc, which favoured a more stable depositional setting with a greater control of the sedimentary processes over the final outcome. Therefore, the geochemical features of the siliciclastic rocks from both domains reveal a change in the tectonic setting over the Ediacaran-Cambrian transition, which preceded the development of a Cambro-Ordovician passive margin.
Sm-Nd isotope composition of the Ediacaran-Cambrian siliciclastic sequences in the Malpica-Tui Complex provides a single population of Nd model ages (1743-2223 Ma) and εNdi values (from -13.1 to -8.1). These rather old Nd model ages and high negative εNdi values observed in the NW Iberian Basal Allochthonous Units account for a dominant contribution from continental crustal sources, which suggests an original proximity to the West African craton during the latest Neoproterozoic and Early Palaeozoic. However, the Ediacaran and Cambrian sequences in the CIZ present εNdi values (-1.7 and -4.7 respectively) and TDM (1288 and 1516 Ma, respectively), which support a general higher contribution from juvenile isotope sources. Moreover slightly older Cambrian Nd isotopic sources suggest greater proximity to the mainland, probably in relation to the suggested widening of the peri-Gondwanan back-arc setting. The contrasting Nd model ages for coeval sedimentary sequences in the NW Iberian Basal Units and the CIZ, likely suggests a different lateral location along the Gondwanan margin. Considering the proximity of the Basal Allochthonous Units to the West Africa craton, the younger Nd model ages observed in the autochthonous CIZ successions place the latter sedimentary basins in a more eastern location, probably in a section of the Gondwana margin located between the West Africa craton and the Sahara Metacraton, with a larger input of siliciclastic material eroded from younger continental source areas.
An Ediacaran origin for the Ecdysozoa?TOP
M. Giacomelli1, R.J. Howard2, J. Lozano-Fernandez1,3, M.A. Wills4, G.D. Edgecombe2, J. Vinther1,2, P.C.J. Donoghue1,3, D.Pisani1,3
1School of Biological Sciences, University of Bristol, UK. Email: davide.pisani@bristol.ac.uk
2The Natural History Museum, London, UK.
3School of Earth Sciences, University of Bristol, UK.
4Department of Biology and Biochemistry, The University of Bath, UK.
Ecdysozoa unites eight phyla of protostome animals: Priapulida, Kinorhyncha, Loricifera (Scalidophora); Nematoda, Nematomorpha (Nematoida); Tardigrada, Onychophora and Euarthropoda (Panarthropoda). Ecdysozoa includes the most diverse and abundant animal phyla (Nematoda and Arthropoda), as well as depauperate lineages (e.g. Priapulida). The ecdysozoan fossil record extends to the early Cambrian, but the precise timing for the origin of this lineage is still debated. To investigate ecdysozoan origins, we assembled a large phylogenomic dataset (up to 228 genes, 66 ecdysozoan and 14 outgrip taxa). For each major ecdysozoan linage, taxa sapling was designed to allow dating the basal split within that lineage. We then applied Bayesian methods to infer a time tree calibrated using a set of 55 fossils. We explored different analytical conditions (e.g. the impact of different calibration strategies and different assumptions about the rate of evolution). Our results corroborate an Ediacaran origin of Ecdysozoa and many major constituent lineages (Scalidophora, Nematoida+Panarthropoda, Panarthropoda and Euarthropoda). We found Nematoida to cross the Cambrian-Ecdysozoan boundary, while Onychophora, Nematoda and Nematomorpha emerged to be Phanerozoic in age.
The Ediacaran-Cambrian Chapel Island Formation of Newfoundland revisited: Evaluating changes in ecospace utilization by early animalsTOP
R.C. Gougeon1, M.G. Mángano1, L.A. Buatois1, G.M. Narbonne1,2, B.A. Laing1
1Department of Geological Sciences, University of Saskatchewan, Saskatoon, SK, S7N 5E2, Canada. Email: gougeon.romain@gmail.com
2Department of Geological Sciences and Geological Engineering, Queen’s University, Kingston, ON, K7L 3N6, Canada.
The Cambrian GSSP (ca. 540 Ma) is marked by the appearance of complex trace fossil from the Treptichnus pedum Ichnofossil Assemblage Zone in the Chapel Island Formation (CIF) at Fortune Head, Newfoundland in Canada (Brasier et al., 1994). The CIF is a 1000 m-thick, most-ly continuous siliciclastic succession that ranges from the late Ediacaran to Cambrian Stage 2. Our present study has relogged this section in detail using the most up to date ichnotaxonomy and facies interpretation. Ichnologic work also focused on reporting bioturbation index (BI; sensu Taylor & Goldring, 1993) and bedding plane bioturbation index (BPBI; sensu Miller & Smail, 1997) and measuring trace fossil width and depth. Tiering and ecospace utilization were evaluated based on this data. Similar depositional environments reoccur through the Fortunian and Cambrian Stage 2 intervals of the CIF, allowing comparisons of ichnofaunas formed in similar environmental settings at different times.
The Fortunian is then characterised by surficial to shallow-tier, diverse assemblages, whereas the Cambrian Stage 2 displays shallow- to deep-tier, moderately to poorly diverse assemblages. Ichnologic analysis through the CIF indicates a temporal increase in burrow size, BI, BPBI and depth of penetration (Gougeon et al., 2018a, 2019). Change in ecospace utilization results from the ecologic engineering effects related to the appearance of large sediment bulldozers and deep-tier suspension feeders during the Fortunian-Cambrian Stage 2 transition (Mángano & Buatois, 2014; Gougeon et al., 2018a, 2019). The bioturbation by deposit feeders and the bioadvective effects of suspension feeders significantly altered the benthic habitat, causing modifications of the sediment structure, sediment stability and increased pore water and solute movements deeper within the sediment.
Loops, circles, spirals and the appearance of guided behaviors from the Ediacaran-Cambrian of Brittany, NW FranceTOP
R. Gougeon1,2, D. Néraudeau2, M. Poujol2, A. Loi3
1Department of Geological Sciences, University of Saskatchewan, Saskatoon, SK, S7N 5E2, Canada. Email: gougeon.romain@gmail.com
2Univ Rennes, CNRS, Géosciences Rennes - UMR 6118, F-35000 Rennes, France.
3Dipartimento di Scienze Chimiche e Geologiche, Università di Cagliari, Cittadella Universitaria, 09042 Monserrato, Italy.
The Ediacaran-Cambrian transition is the place of striking changes in Earth ecosystems, with a diversification of life recorded by a complexification in animal behaviors. In a series of classic papers, Crimes (1974, 1987, 1992a, b) compiled worldwide data on trace fossil distribution from that time period, evaluating their potential as biomarkers and deciphering evolutionary processes. He suspected that many trace fossils typical of Phanerozoic deep-water settings originated in shallower environments. Spiraling is a strategy developed by organisms to survive in environments depleted in nutrient, exemplified by the farming open-burrow system Spirorhaphe (Seilacher, 1977). On the other hand, simple horizontal spirals of deposit-feeders are represented by many ichnogenera, common forms being Spirodesmos and Spirophycus (Buatois et al., 2017).
Spiral trace fossils are poorly reported from the Ediacaran-Cambrian transition and remain problematic (e.g. Jensen, 2003; Carbone & Narbonne, 2014). Conversely, simple looping horizontal trails, fully circling (i.e. Circulichnis) or self-overcrossing (i.e. Gordia) are easily and abundantly identified. Crimes (1992a) suspected spiral trace fossils to appear stratigraphically higher than simple looping trails, at the same level of treptichnids (n.b. it is interesting to note that the two spiralling traces at issue, Planispiralichnus and Protospiralichnus, are omitted in another list in Crimes, 1992b; these Russian forms are now suspected to represent circular treptichnids by Marusin, 2011).
The Brioverian from central Brittany, NW France, is a thick siliciclastic sedimentary succession that has been intensely deformed due to the successive Cadomian and Variscan orogenetic events. Hence, its stratigraphy, basin architecture and fossil content have been poorly understood, and the position of its uppermost limit within the Ediacaran or the Cambrian is still a pending question. However, recent investigations reported insights of life in the form of simple trace and body fossils in shallow-marine, tidally influenced settings (Gougeon et al., 2018b; Néraudeau et al., 2018). Following up this promising start, new investigations were conducted and new ichnofossils were reported from outcrops in Saint-Gonlay, Montfort-sur-Meu (eastern Brittany) and Crozon (western Brittany; Fig. 3). From these, abundant horizontal looping, fully circling and pseudo- to perfect-spiraling trails are found associated with microbially stabilized surfaces. Although the two first forms can be identified as Gordia and Circulichnis respectively, the ichnotaxonomic position of the latest ones is more uncertain. Regardless of their actual name, spiral trails are not uncommon in the section and show that more complex programs (i.e. phobotaxis and thigmotaxis) were already acquired by their tracemakers.
|
 Figure 3.—Looping, circling and spiraling trace fossils from the Brioverian. Scale bars are 1 cm. 1. Gordia with numerous self-overcrossings. Montfort-sur-Meu (les Grippeaux). 2. Circular Gordia with one self-overcrossing (arrow). Saint-Gonlay (la Lammerais). 3. Tiny Circulichnis on a microbially stabilized surface. Note that the trail starts (or ends) outside of the circle (arrow). Saint-Gonlay (la Lammerais). 4. Perfect tiny Circulichnis (top right) associated with horizontal trails (left) and a pit of indistinct affinity (bottom right). Saint-Gonlay (la Lammerais). 5. Pseudo-spiraling Gordia with a self-overcrossing (arrow) and an unguided course. Montfort-sur-Meu (le Bois-du-Buisson). 6. Spiral trail with an uncertain guided course. Montfort-sur-Meu (le Bois-du-Buisson). 7-8. Perfect spiral trails with guided courses. Montfort-sur-Meu (les Grippeaux; 7) and Crozon (8). Figure 3.—Looping, circling and spiraling trace fossils from the Brioverian. Scale bars are 1 cm. 1. Gordia with numerous self-overcrossings. Montfort-sur-Meu (les Grippeaux). 2. Circular Gordia with one self-overcrossing (arrow). Saint-Gonlay (la Lammerais). 3. Tiny Circulichnis on a microbially stabilized surface. Note that the trail starts (or ends) outside of the circle (arrow). Saint-Gonlay (la Lammerais). 4. Perfect tiny Circulichnis (top right) associated with horizontal trails (left) and a pit of indistinct affinity (bottom right). Saint-Gonlay (la Lammerais). 5. Pseudo-spiraling Gordia with a self-overcrossing (arrow) and an unguided course. Montfort-sur-Meu (le Bois-du-Buisson). 6. Spiral trail with an uncertain guided course. Montfort-sur-Meu (le Bois-du-Buisson). 7-8. Perfect spiral trails with guided courses. Montfort-sur-Meu (les Grippeaux; 7) and Crozon (8).
|
|
An Ediacaran double spiral from Australia closely relates in shape and size to the holotype of Spirorhaphe involute (Jensen, 2003). This trace fossil is transitional with unguided meanders, and the preservation as a trail or burrow is unknown; an affinity with a farming open-burrow system is then difficult to conceive. Beside that report, the oldest true Spirorhaphe comes from the Ordovician (Pickerill, 1980; Lehane & Ekdale, 2016). Regarding simple deposit-feeder spirals, they are reported from the Ediacaran-Cambrian transition in both shallow- (this study) and deep-marine settings (Carbone & Narbonne, 2014). Microbially stabilized surfaces were pervasive at that time and may have been a wealthy medium for the nutrition of trail producers. Increasing sediment processing by penetrative animals during the Cambrian resulted in the progressive retreat of microbial mats in deep-marine settings (Crimes, 1992a, b; Buatois et al., 2011). After their disappearance by the Ordovician, eventually, organisms adapted their simple spirals of deposit-feeders to these nutrient-poor environments by developing more complex, farming open-burrow systems.
Ediacaran origins of complex animal behaviour: trace fossil evidence from the Hoogland Formation of NamibiaTOP
J.F. Hoyal Cuthill1,2, C. Kenchington2, T. Boag3, M. Laflamme4
1Institute for Analytics and Data Science and School of Life Sciences, University of Essex, Wivenhoe Park, Colchester, CO4 3SQ. Email: j.hoyal.cuthill@elsi.jp
2Department of Earth Sciences, University of Cambridge, Downing Street, Cambridge, CB2 3EQ, UK.
3Department of Geological Sciences, University of Stanford, Braun Hall (Geo Corner) #118, 450 Serra Mall, Building 320, Stanford, CA 94305-2115.
4Department of Chemical and Physical Sciences, University of Toronto, 359 Mississauga Road, Mississauga, Ontario, L5L 1C6, Canada
Trace fossils of subsurface burrowing provide the earliest evidence for bilaterian animals, mesodermal musculature, the through gut, a centralised nervous system, and complex behaviour (Brasier et al., 2013). Definitive body fossils of bilaterian animals appear during the Cambrian ‘Explosion’ of animal diversity (541-485 Ma). However, the record of characteristically bilaterian activity, including subsurface burrowing, is now known to extend below this, to the base of the Cambrian at 541 Ma and into the latest Ediacaran Period (635-541 Ma) (Han et al., 2004; Vannier et al., 2010; Parry et al., 2017). The global stratotype section and point (GSSP) at the base of the Cambrian is defined by the first appearance of the trace fossil Treptichnus pedum (previously called Phycodes pedum) (Brasier et al., 1994; Vannier et al., 2010). This is a subhorizontal, segmented burrow system (Vannier et al., 2010) originally suggested to be markedly more complex than earlier, Ediacaran trace fossils (Brasier, et al., 1994). However, Treptichnus is now known from the latest Ediacaran of Newfoundland and Namibia (Vannier et al., 2010). Furthermore, the complex trace fossil Streptichnus narbonnei, with apparent similarities to Treptichnus, has been described from the Spitskopf Member of the Urusis Formation in Namibia, of terminal Ediacaran or lowermost Cambrian age (Jensen & Runnegar, 2005). This suggests that the origins of complex animal behaviour could significantly predate the Cambrian record of bilaterian diversification. However, complex trace fossils from Ediacaran rocks have thus far been found relatively close to the Cambrian boundary and have not been subjected to quantitative analysis of behavioural complexity (Sims et al., 2014). Here we describe quantitatively a new segmented trace fossil, Streptichnus vagans ich. sp. nov., represented by a unique specimen stratigraphically located from the Hoogland Member of the Zaris Formation, Zebra River, Namibia, which has been previously radiometrically dated to 547.32 ± 0.65 (Gradstein, 2012). S. vagans shows similarities to the complex, segmented feeding trace fossils T. pedum and S. narbonnei but also features distinct from those of previously described ichnotaxa. Distinct features of S. vagans include the combination of a segmented structure, shallowly multidirectional path (e.g. differing from the unidirectional, clustered S. narbonnei) and consistent segment orientation across multidirectional turns (e.g. differing from T. pedum). S. vagans therefore represents one of the earliest known examples of a “complex” burrowing trace fossil, pushing back the origins of complex animal behaviour to approximately six million years before the beginning of the Cambrian (dated to 541 Ma, Gradstein 2012). Statistical analyses of the turning angle distribution for S. vagans, measured between successive segment centres, show a non-normally, positively skewed distribution, which is typical of optimal animal foraging behaviour in heterogeneous environments with sparse or patchy resources (Sims et al., 2014). This provides a new snapshot of the behavioural and ecological conditions of early animal complexity, showing a lone wanderer exploring a sparse resource landscape.
Constraining morphological disparity in RangeomorphsTOP
C.G. Kenchington1, F.S. Dunn2,3, A.G. Liu1, P.R. Wilby3
1Department of Earth Sciences, University of Cambridge, Downing Street, Cambridge, CB2 3EQ, UK. Email: kenchinc@tcd.ie
2Oxford University Museum of Natural History, University of Oxford, Parks Road, Oxford, OX1 3PW, UK.
3British Geological Survey, Nicker Hill, Keyworth, Nottingham, NG12 5GG, UK.
Avalonian Ediacaran successions (those from Newfoundland, Canada, and Charnwood Forest, UK; c. 575-560 Ma) host the oldest known communities of complex multicellular organisms, including some of the best candidate early animals. Rangeomorphs dominate these communities, and are the most geographically and environmentally widespread of the Ediacaran clades. Although the gross morphology of rangeomorph taxa is broadly similar, some species bear morphological structures that are unique among these groups. For example, the recently discovered “brushes” from Charnwood Forest bear a unique external sheath-like structure, while some multifoliate taxa, including Hylaecullulus fordi, demonstrate novel eccentric growth architecture.
Diversity and species richness are critical parameters in studies of modern and fossil ecology. However, where these measures are influenced by changing opinions on classification, morphological disparity is not. Here we present an analysis of the disparity of rangeomorph taxa, across 20 communities, using metric and non-metric dimensional scaling and multivariate analysis. Our analyses are coded to include intraspecific variation, which has recently been recognised to be considerable in some rangeomorphs taxa. Our disparity framework provides a tool with which we then test how species with unique features (e.g. the “brushes”) fit within the rest of the known rangeomorph taxa. Finally, quantifying morphological disparity in rangeomorphs allows their levels of disparity to be compared to that of extant groups, and provides a quantitative assessment of the biotic composition of different communities.
Tracking the diversification of the Ediacaran macrobiota: Tying fossil occurrence data to a robust chronostratigraphic framework at Mistaken Point UNESCO World Heritage SiteTOP
A.G. Liu1, J.J. Matthews2,3, B. Levell4, C. Yang5, D. McIlroy2, D.J. Condon5
1Department of Earth Sciences, University of Cambridge, Downing Street, Cambridge, CB2 3EQ, UK. Email: agscl2@cam.ac.uk
2Department of Earth Sciences, Memorial University of Newfoundland, 300 Prince Philip Drove, St. John’s, NL, A1C 3X5, Canada.
3Oxford University Museum of Natural History, Parks Road, Oxford, OX1 3PW, UK.
4Department of Earth Science, University of Oxford, South Parks Road, Oxford, OX1 3AN, UK.
5British Geological Survey, NERC Isotope Geosciences Facilities, Nicker Hill, Keyworth, NG12 5GG, UK.
Mistaken Point Ecological Reserve UNESCO World Heritage Site (MPER) is an internationally recognised locality for fossils of the Ediacaran macrobiota, hosting the oldest known examples of these organisms, including candidate metazoan body and trace fossils. Recent construction of a chronostratigraphic framework for the Conception and St. John’s Groups sedimentary succession within MPER (see Matthews et al. abstract for this meeting) offers an unparalleled opportunity to constrain the tempo of the rise and diversification of the MPER biotas.
Here we present a compilation of fossil occurrence data spanning 80 discrete fossil-bearing horizons within the MPER succession. These data yield stratigraphic ranges for all formally described macrofossil taxa within the MPER succession, facilitating recognition and interrogation of potential evolutionary signals. Plotting these data against both lithostratigraphy and time reveals likely facies controls on the occurrence of certain taxa, and provides temporal constraint on the appearance of key groups.
Peak taxonomic diversity is recognised within the upper Mistaken Point and lower Trepassey Formations. Heterogeneity in both population and species abundance is observed between and within palaeocommunities. Uniterminal rangeomorphs appear within the MPER stratigraphic succession several million years before multiterminal forms. Comparison of our occurrence data with similar stratigraphic range data we have collected from the Bonavista Peninsula (~200km to the NW) provides support to the suggestion that the MPER patterns are real evolutionary signals.
Together, our combined stratigraphic, palaeontological and geochronological approach offers a holistic record of evolution during the mid-late Ediacaran Period, and enables the development of testable hypotheses regarding the evolutionary trajectory of rangeomorphs and associated Avalonian taxa.
A Nenoxites-Horodyskia fossil assemblage from the Liuchapo Formation, Liujiata village, Hunan Province, China: a glimpse at the Ediacaran-Cambrian life in deep-water settingsTOP
C. Luo1, L. Miao2
1CAS Key Laboratory of Economic Stratigraphy and Palaeogeography, Nanjing Institute of Geology and Palaeontology and Center for Excellence in Life and Paleoenvironment, Chinese Academy of Sciences, 39 East Beijing Road, Nanjing 210008, China. Email: cluo@nigpas.ac.cn
2State Key Laboratory of Palaeobiology and Stratigraphy, Nanjing Institute of Geology and Palaeontology and Center for Excellence in Life and Paleoenvironment, Chinese Academy of Sciences, 39 East Beijing Road, Nanjing 210008, China.s
So far, our understanding of the life evolution crossing the Ediacaran-Cambrian interval was mainly based on fossil assemblages from platform and inner-ramp facies, while much less is known from deeper facies such as slopes and basins. Here a fossil assemblage is described from the chert of the Liuchapo Formation in Liujiata Village, Hunan Province, which represents a slope-to-basinal setting at the terminal Ediacaran and earliest Cambrian.
The base of the outcrop starts with 10 m of dolostones intercalated with thin chert layers and chert nodules (Dengying Formation). This is overlain by an approximately 50 m chert succession (Liuchapo Formation), which is only interfered by shale and dolostone beds at around 30 m above the Dengying-Liuchapo contact (thereafter abbreviated as DLB). At the top of the Liuchapo Formation, chert gradually turns to the silicified black shale of the Niutitang Formation. According to a previous U-Pb radiometric dating of tuffaceous beds from an adjacent correlative outcrop, the base of the Liuchapo Formation is only slightly older than 541.0 ± 1.0 Ma (Chen et al., 2015).
The cherty layers and nodules in the Dengying Formation and the whole Liuchapo Formation were sampled and analyzed. Despite fragmented organic remains from the chert layers of the Dengying Formation, unequivocal Horodyskia appears at 16 m above DLB. Nenoxites probably first appears in the dolostone at 30 m above DLB and is unambiguously preserved in the chert just above this level. The fossil assemblage increases in abundance and diversity upwards and reaches a climax at around 49-57 m. In this interval, Nenoxites prevails over Horodyskia, and some enigmatic tubular fossils of different sizes are also preserved. Further above, the fossil abundance and diversity decrease rapidly. The last unambiguous member of the assemblage, Horodyskia, disappears above 58 m.
Horodyskia-Nenoxites assemblages have been previously reported from slope-basin cherts of the Liuchapo and Piyuancun Formations from Guizhou and Anhui Provinces (Dong et al., 2008, 2012). A condensed distribution of Nenoxites was also described from offshore carbonate settings from Siberia (Rogov et al., 2012). However, the intimate co-occurrence of all the different sorts of fossils, the enigmatic tubular forms, and the vertical change of the abundance and diversity of the assemblage were not been described elsewhere.
By studying the structure and preservation of the massively preserved fossils using optical microscope, SEM and Raman spectroscopy, we found that Nenoxites, as well as the Horodyskia and the enigmatic tubular fossils, are probably not animal traces as some researchers suggested (e.g. Rogov et al., 2012), because:
(i) In the fabric, where multiple sorts of fossils are densely packed, the fossils never cross each other. Interruption even never happens between different sorts of fossils.
(ii) All fossils are preserved with clay minerals in different types of cherts and dolostones, while their analogues in siliciclastic and muddy limestone were also reported to be preserved in such a composition. This is hardly a character of trace fossils, in which the burrow infill is expected to be varied in different sedimentary facies.
(iii) Incongruent with the backfilling model, the sediments in the interspaces between the bowl-shaped units of Nenoxites show no difference from the host rock. Furthermore, it has also been observed in our material that a Horodyskia bead fell in a bowl-shaped unit of Nenoxites.
(iv) The investigated fossil assemblage is located in a slope-basinal setting. According to the current oxygenation models for the Ediacaran-Cambrian ocean, the distal deep water was generally euxinic and ferruginous (e.g. Li et al., 2018). Although there may have been some the transiently oxygenated intervals, the oxygen level could only support some sessile animals like sponges (e.g., Cheng et al., 2017). If this model is correct, such an environment could hardly allow intensive animal burrowing.
For these reasons, we interpret Horodyskia, Nenoxites and the enigmatic tsubular fossils as probable eukaryotic organisms which have an unknown intrinsic character to trigger an early diagenetic, post-mortem mineralization of clay minerals. In light of the interpretation, the massively preserved fossils in this study, and those reported from Siberia, likely represent thick “mats” of these organisms. In non-laminated cherts, we did find structures resembling EPS shrinkage in microbial mats. However, further investigations are under way to determine whether the fossil assemblage is laterally correlative and synchronous in a larger area, and more geochemical analyses are to be employed to reveal the possible metabolic mechanism of these organisms.
Ichnotaxonomic conundrums and the calibration of the Cambrian explosion: The case of Psammichnites-TaphrhelminthopsisTOP
M.G. Mángano1, L.A. Buatois1, S. Jensen2, R. MacNaughton3, A. Marcos4, L. Piñuela5, J.C. García-Ramos5, R. Gougeon1
1Department of Geological Sciences, University of Saskatchewan, 114 Science Place, Saskatoon, Saskatchewan, S7N 5E2, Canada. Email: gabriela.mangano@usask.ca, luis.buatois@usask.ca, gougeon.romain@gmail.com
2Área de Paleontología, Facultad de Ciencias, Universidad de Extremadura, E-06006, Badajoz, Spain. Email: soren@unex.es
3Geological Survey of Canada (Calgary), Natural Resources Canada, Calgary, Alberta T2L 2A7, Canada. Email: robert.macnaughton@canada.ca
4Departamento de Geología, Universidad de Oviedo, c/ Jesús Arias de Velasco s/n 33005 Oviedo, Asturias, Spain. Email: marcos@geol.uniovi.es
5Museo del Jurásico de Asturias (MUJA), 33328 Colunga, Asturias, Spain. Email: lpinuela.muja@gmail.com, jcgramos.muja@gmail.com
Assessing changes in ichnodiversity through time is becoming an important tool to calibrate evolutionary radiations. However, the soundness of these studies necessarily depends on a well-supported ichnotaxonomy. In particular, reconstructing ichnodiversity trajectories during the Cambrian explosion is based on compilations of global datasets, which in turn involve multiple ichnotaxonomic decisions, such as careful evaluation of preservational variants and assessment of potential synonyms. In addition, the contrasting philosophical approaches of splitters and lumpers permeate any taxonomic work.
The taxonomic status of Taphrhelminthopsis circularis, an ichnospecies originally described from lower Cambrian shallow-marine deposits of Asturias, Spain (Crimes et al., 1977), illustrates this problem. The ichnogenus Taphrhelminthopsis was originally introduced for trace fossils with a bilobate lower surface recorded in post-Paleozoic deep-marine deposits. However, it has been convincingly argued that it represents a preservational variant of Scolicia (Uchman, 1995). Notably, Taphrhelminthopsis has also been profusely used for Cambrian shallow-marine forms, whose ichnotaxonomic affinity remains elusive. Taphrhelminthopsis circularis was originally defined as consisting of bilobate trace fossils with a central furrow and a tendency to form a circular course. Taphrhelminthopsis circularis has been subsequently recorded in many lower Cambrian strata worldwide. The fact that no information on toponomy was provided in its original description has complicated a critical re-evaluation of this ichnotaxon. Re-analysis of the type specimen and study of additional material at the type locality demonstrate that T. circularis occurs at the top of sandstone layers, with an upward convex bilobate morphology, confirming the suspicion that it could represent a preservational variant of Psammichnites. In addition, the validity of the ichnospecies is questioned based on the variable course recorded by topotype specimens. Affinities with Psammichnites are also inferred for other trace fossils commonly recorded from Cambrian shallow-marine strata under a wide variety of names, such has Taphrhelminthopsis nelsoni, Taphrhelminthoida dailyi, Taphrhelminthoida ?convoluta, Helminthoida crassa and H. miocenica. (e.g. Hofmann & Patel, 1989).
Documenting the multiple preservational variants of Cambrian Psammichnites is important in order to calibrate ichnodiversity levels and evaluate the effects of novel styles of animal-substrate interactions during the Cambrian explosion. Global compilations indicate a dramatic ichnodiversity increase during the early Cambrian, which is consistent with the notion of the Cambrian explosion. The recent suggestion that this increase may be an artifact resulting of counting preservational variants (i.e. taphomorphs) of the same ichnotaxon as different ichnotaxa is unsupported because the available datasets have actually filtered these potential taphonomic variants.
Brachiopod-based biostratigraphy of the Kaili Formation, Cambrian Series 2, Stage 4 to Miaolingian Series, Wuliuan Stage at Balang, Jianhe, Guizhou, South ChinaTOP
Y.Q. Mao1, Y.L. Zhao2, Y.J. Liu2
1Institute of Mountain Resources, Guizhou Academy of Sciences, 550001 Guiyang, Guizhou Province, PR China.
2College of Resources and Environmental Engineering, Guizhou University, 550025 Guiyang, Guizhou Province, PR China. Email: zhaoyuanlong@126.com
The Wuliu-Zengjiayan section, global standard stratotype Section of Cambrian Miaolingian Series and Wuliuan Stage, yields abundant trilobites, acritarchs and brachiopods in Jianhe County, Guizhou Province. Brachiopod fossils are assigned to two subphyla, containing 11 genera and 13 species. The brachiopods are mainly distributed in the lower, middle and upper parts of the Kaili formation at the Wuliu-Zengjiayan section, especially in the middle part. Based on their stratigraphic ranges, the Kaili Formation is divided into three brachiopod assemblages, from bottom to top (Fig. 1).
1. Nisusia guizhouensis-Palaeobolus bretonensis Assemblage (Figs. 4-5, 6B, E, G). This assemblage of two species corresponds to the Bathynotus kueichouensis-Ovatoryctocara sinensis Assemblage Zone of the Kaili Formation.
2. Paterina jianhensis-Kutorgina sp. Assemblage (Figs. 4-5, 6A-C), most fossiliferous, which includes Paterina jianhensis, Kutorgina sp., Lingulella bynguanoensis, Lingulepis taijiangensis, Linnarssonia tumida, Dictyonina sp., Micromitra sp. and Eoconcha sp., and corresponds to the Oryctocephalus indicus Zone, the middle trilobite zone of the Kaili Formation.
3. Acrothele bohemica-Acrothele kailiensis sp. Assemblage (Figs. 4-5, 6D-E) includes three species, Linnarssonia tumida, Acrothele bohemica and Acrothele sp., which corresponds to the Peronopsis taijianggesis Zone, the upper trilobite zone of the Kaili Formation.
|
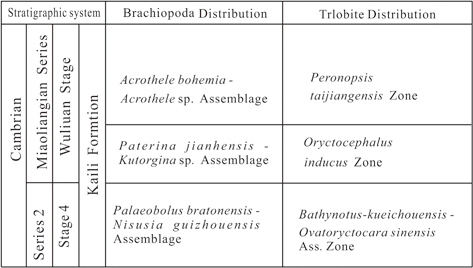 Figure 4.—Correlation chart of the brachiopod assemblages and the trilobite zones of the Kaili Formation at Balang, Jianhe, Guizhou, South China; modified from Mao (2017), Mao et al. (2017), Zhao et al. (2017) and Peng & Zhao (2018). Figure 4.—Correlation chart of the brachiopod assemblages and the trilobite zones of the Kaili Formation at Balang, Jianhe, Guizhou, South China; modified from Mao (2017), Mao et al. (2017), Zhao et al. (2017) and Peng & Zhao (2018).
|
|
|
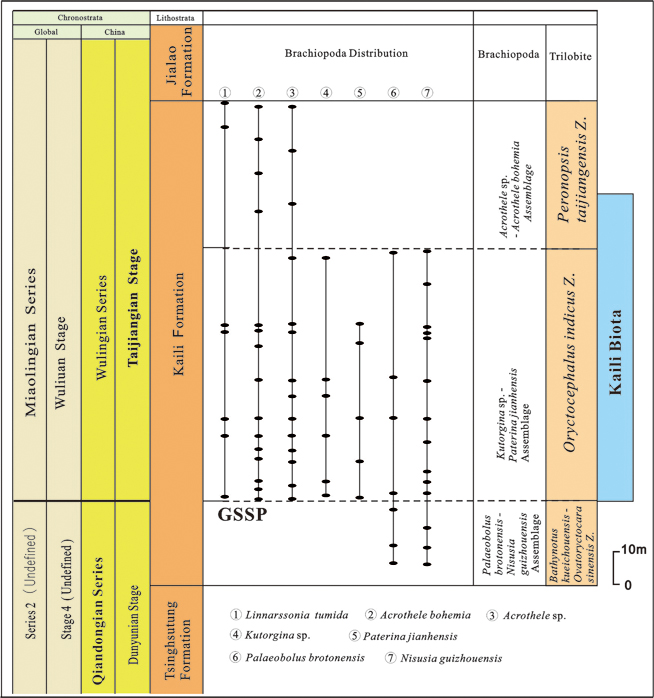 Figure 5.—Stratigraphic distribution of brachiopods in the Kaili Formation at Wuliu-Zengjiayan section, China. Figure 5.—Stratigraphic distribution of brachiopods in the Kaili Formation at Wuliu-Zengjiayan section, China.
|
|
|
 Figure 6.—Specimens collected from the Kaili Formation in Balang village, Jianhe, Guizhou, South Chnia. Scale bars = 2 mm, except F = 5 mm (A-E, G). A. Kutorgina sp. (GTBJ-15-3), external view of ventral valve. B. Nisusia guizhouensis (GTBJ-20-3), external view of ventral valve. C. Paterina jianhensis (GTBM-10-1502), external view of ventral valve. D. Acrothele sp. (GTBJ-9-2-3675), external view of dorsal valve and ventral valve. E. Acrothele bohemica (GTBM-9-3-5181), external view of dorsal valve and ventral valve. F. Palaeobolus bretonensis (GTMJ-12-37), external view of dorsal valve. G. Nisusia guizhouensis (GTBJ-16-106), external view of dorsal valve. Figure 6.—Specimens collected from the Kaili Formation in Balang village, Jianhe, Guizhou, South Chnia. Scale bars = 2 mm, except F = 5 mm (A-E, G). A. Kutorgina sp. (GTBJ-15-3), external view of ventral valve. B. Nisusia guizhouensis (GTBJ-20-3), external view of ventral valve. C. Paterina jianhensis (GTBM-10-1502), external view of ventral valve. D. Acrothele sp. (GTBJ-9-2-3675), external view of dorsal valve and ventral valve. E. Acrothele bohemica (GTBM-9-3-5181), external view of dorsal valve and ventral valve. F. Palaeobolus bretonensis (GTMJ-12-37), external view of dorsal valve. G. Nisusia guizhouensis (GTBJ-16-106), external view of dorsal valve.
|
|
The division and establishment of the stratigraphic ranges of the brachiopods included in the Kaili Formation at the Wuliu-Zengjiayan section, global standard stratotype Section of the Cambrian Miaolingian Series and Wuliuan Stage, is an essential supplement to the biostratigraphic division of trilobites and acritarchs.
This study was supported by the Ministry of Science and Technology of China (Grant No.41772021 to Yang, Zhao)
Sinotubulites growth and relationship with CloudinaTOP
M. Martí Mus1, I. Cortijo2, T. Palacios1
1Área de Paleontología, Facultad de Ciencias, Universidad de Extremadura, 06006 Badajoz, Spain. Email: martimus@unex.es
2Villuercas-Ibores-Jara UNESCO Global Geopark, Pintores 10, 10003 Cáceres, Spain.
The first biomineralizing metazoans inhabited late Ediacaran carbonate settings and represent a relatively diverse assemblage, including at least tree genera: Cloudina, Sinotubilites and Namacalathus. While several similarities have been noted, particularly between Cloudina and Sinotubulites, the phylogenetic relationship between these early biomineralizers is unclear, as is their position in the metazoan phylogenetic tree. Also unknown are many aspects of tsheir physiology and ecology, including their detailed mode of growth (only partly known for Cloudina) and the timing of the mineralization process.
Sinotubulites is a millimetric to centimetric, multilayered tube which often has an irregular polygonal cross section. Its detailed morphology is variable and depends on tube ornamentation. There are two distinct “blocks” of layers in the Sinotubulites tube: an outer block formed by irregularly wrinkled layers, which give the tube its somewhat polygonal outline, and an inner block formed by smooth, cylindrical layers. In the outer block, the degree of ornamentation increases towards the exterior, and the ornamentation pattern is somewhat replicated and augmented by successive layers. Specimens described to date are open at both ends, but are likely fragmentary. As is the case with the co-occurring Cloudina, folding patterns suggest the layers were secreted unmineralized. There is no evidence of growth lines in the skeletons of Cloudina or Sinotubulites, and all available data suggests that each funnel of Cloudina was secreted “at once”, as a cuticle of pliable organic matter shaped by the soft parts. Both in Cloudina and Sinotubulites, occasional fractures of the wall are interpreted as indicative of mineralization, and the presence and distribution of Sr suggest an original aragonitic mineralization.
Material from the Ibor Group, Villarta de los Montes, central Spain, sheds light on the mode of growth of Sinotubulites, and evidences its close phylogenetic relationship with Cloudina. Sinotubulites grew in a similar way to Cloudina, by progressively adding “funnel”-shaped growth units terminally. However in the case of Sinotubulites each “funnel” bends over itself, embracing the anterior portion of the tube. The growth unit of Sinotubulites forms therefore a pair of continuous, concentric cylinders. The internal cylinder is smooth and must have been secreted by the soft parts occupying the lumen of the tube. The addition of successive smooth cylinders, from the inside of the tube, forms the internal block of layers of the Sinotubulites tube. Proximally, the smooth cylinder flares and bends over itself forming a second, irregularly folded, embracing layer, which is added, this time from the outside, to the external block of layers. Each new external layer accommodates to the previous one, enhancing its ornamentation. This construction implies that, at least during each growth phase, the soft parts of the Sinotubulites animal must have embraced the whole proximal portion of the tube, secreting and moulding the new cuticle, pressing it against the previous one.
It has been assumed that both Cloudina and Sinotubulites represent external skeletons, once inhabited by a worm- or polyp-like small metazoan. However the data presented herein shows that, at least during each growth phase, the soft parts of Sinotubulites extended well beyond the tube aperture, and must have been relatively large, perhaps unable to withdraw entirely in the relatively narrow central lumen. This in turn suggests that the function of the skeleton was not protection but more likely related with substrate colonization and stability in a carbonate setting.
Stratigraphic and sedimentological investigation of the Ediacaran Jodhpur Group in and around Jodhpur city, IndiaTOP
S.C. Mathur1, V.S. Parihar1, H. Ram1, S. Mathur1
1Department of Geology, J.N.Vyas University, Jodhpur, India. Email: sureshsushma09@gmail.com
2Centre for climate change and water research, Suresh Gyan Vihar University, Jaipur, India.
Geologically speaking, the Marwar Supergroup (Neoproterozoic to Cambrian) is a significant entity of the western Rajasthan, India. It has been subdivided into four major groups, from bottom to top: the Jodhpur, Bilara, Hanseran and Nagaur groups (Pareek, 1984; Chauhan et al., 2001). The outcrops of the Jodhpur Group (JG) cropping out in and around Jodhpur city occur at Umaid Palace, Daijar, Mandore, Sursagar, Keroo, Beroo, Keru, Kali Beri, Bhuri Beri, Singoria hill, Sidhnath hill, Gangana and Motisar areas. Stratigraphically, 220 m of JG overlies the Maiani Igneous Suite of rocks (745 to 681 Ma) and underlies the Quaternary Jodhpur city Formation (JCF). JG was previously subdivided into the Pokaran Boulder Bed, Sonia Sandstone and Giribhakar Sandstone, but this arrangement is in need of reevaluation. Based on lithological assemblages, lithological distributions in the field, basement cover relationships, facies analysis, sedimentary structures and their depositional environments, the stratigraphy of JG at Jodhpur is revised below. Accordingly, JG is divided into a conglomerate unit at the base followed by the Umaid Bhawan (UBF), Sursagar (SSF) and Motisar (MF) formations (Figs 7-9).
|
 Figure 7.—Lithostratigraphy of Marwar Super Group showing revised stratigraphy of Jodhpur Group of Jodhpur city (modified from Sarkar et al., 2008). Figure 7.—Lithostratigraphy of Marwar Super Group showing revised stratigraphy of Jodhpur Group of Jodhpur city (modified from Sarkar et al., 2008).
|
|
|
 Figure 8.—Outcrops of the Malani Igneous suite of rocks surrounding Mehrangarh Fort and Jaswant palace (left). B. Contact between the Malani Igneous suite and the Jodhpur Group marked by a basal conglomerate (centre). C. Contact of rhyolite of MIS and Umaid Bhawan Formation of Jodhpur Group (right). Figure 8.—Outcrops of the Malani Igneous suite of rocks surrounding Mehrangarh Fort and Jaswant palace (left). B. Contact between the Malani Igneous suite and the Jodhpur Group marked by a basal conglomerate (centre). C. Contact of rhyolite of MIS and Umaid Bhawan Formation of Jodhpur Group (right).
|
|
|
 Figure 9.—Outcrops of the Sursagar Formation in an open quarry (left). B. Contact between sandstones of the Sursagar Formation (lower) and sandstones of the Motisar Formation (upper) from the Jodhpur Group (centre). C. Wave ripples on sandstones of the Sursagar Formation (right). Figure 9.—Outcrops of the Sursagar Formation in an open quarry (left). B. Contact between sandstones of the Sursagar Formation (lower) and sandstones of the Motisar Formation (upper) from the Jodhpur Group (centre). C. Wave ripples on sandstones of the Sursagar Formation (right).
|
|
All these successions are overlain by the Quaternary JCF. The basal conglomerate consists dominantly of pebbles of rhyolite, subordinate amount of vein quartz, jasper and granite pebbles set in a fine- to medium-grained quartz-felspathic matrix. The clast/matrix ratio suggests that it is ranging from clast- to matrix-supported, oligomictic to rarely polymictic in nature, and the deposits were sedimented as channel lags to fluvial fan-apron deposits. The coarsening upward UBF is represented by claystone at the base followed by silty shale, fine- to medium and coarse-grained sandstone and pebbly sandstone at the top. Dominantly, festoon cross-bedding bearing UBF indicates a deltaic environment. The SF is characterized by fine- and medium-grained sandstones typically of quartz arenitic nature. It shows a prolific development of wavy ripple marks, scour and fill, rib and furrow structures with Ediacaran fauna (Kumar & Pandey, 2009; Sharma & Mathur, 2014; Parihar et al., 2015; Mathur et al., 2017), representing typical near coastal to beach deposits. MF consists of eight cycles of fining-upward sequences including a conglomerate grading upwards into a pebbly, medium- and fine-grained sandstone. MHF is characterized by the presence of large-scale trough cross-beds dominantly denoting fluvial environments. The sediments of JCF comprise clayey, silty to sandy alluvial deposits sedimented in the Jodhpur valley areas, and represent fluvial environments. All formations of JG, at one or other places, directly rest on the MIS basement. Secondly, the occurrence of sandstone tablets resting on rhyolite at more than 260 m height as well as at the ground level are stratigraphically puzzling. It has been found that the distribution and deposition of sediments of JG are governed by the configuration of basement rocks and their subsequent sedimentological processes. The paper embodies stratigraphic and sedimentological analysis to understand the facies distribution and stratigraphic setup of Ediacaran fauna bearing JG of Marwar Supergroup in and around the Jodhpur city.
Constraining the rise of the Ediacaran biota: A new chronostratigraphic framework from the Mistaken Point UNESCO World Heritage SiteTOP
J.J. Matthews1,2, D.J. Condon3, B. Levell4, C. Yang3, D. McIlroy1, A.G. Liu5
1Department of Earth Sciences, Memorial University of Newfoundland, 300 Prince Philip Drove, St. John’s, NL, A1C 3X5, Canada. Email: jjmatthews@mun.ca
2Oxford University Museum of Natural History, Parks Road, Oxford, OX1 3PW, UK.
3British Geological Survey, NERC Isotope Geosciences Facilities, Nicker Hill, Keyworth, NG12 5GG, UK.
4Department of Earth Science, University of Oxford, South Parks Road, Oxford, OX1 3AN, UK.
5Department of Earth Sciences, University of Cambridge, Downing Street, Cambridge, CB2 3EQ, UK.
The Conception and St. John’s Groups of south eastern Newfoundland, Canada, contain some of the oldest known examples of the Ediacaran macrobiota. Mistaken Point Ecological Reserve UNESCO World Heritage Site (MPER) is internationally recognised for such fossils, hosting both the oldest known palaeocommunities of large, architecturally complex life anywhere in the world, and the oldest evidence of metazoan-style locomotion. Understanding the causes of the emergence, evolution and extinction of the Ediacaran macrobiota, and their relationship to environmental change, requires the construction of an accurate, highly-resolved chrono-stratigraphic framework upon which biostratigraphic, taphonomic, sedimentological, and environmental data can be integrated. The MPER sedimentary succession, including more than 1000m of fossil-bearing strata alongside numerous datable tuffaceous horizons, is ideally suited to this endeavour.
We here present six U-Pb ID-TIMS radio-isotopic ages derived from zircons within volcaniclastic beds of the Conception and St. John’s Groups at MPER. Detailed sedimentological analysis reveals the depositional processes associated with the dated tuffites, contextualising our understanding of both the ages yielded, and the taphonomy of associated Ediacaran macrofossils. Remapping of the MPER stratigraphic succession enables us to present an updated lithostratigraphy in which to place these new ages.
Our data provide high-precision age constraints on horizons including the oldest architecturally complex macrofossils in the MPER succession from the upper Drook Formation; the youngest MPER rangeomorph fossils from the Fermeuse Formation; the famously diverse fossil assemblage of the ‘E’ Surface; and exceptionally preserved specimens on the ‘Brasier’ Surface in the Briscal Formation. We compare and contrast our ages to previously published datasets, and use our new ages to construct an age model for the MPER sedimentary succession, that is consistent with deposition within a back-arc basin. The chronostratigraphic framework we present permits the evaluation of regional palaeobiological datasets relative to both lithostratigraphy and time, broadening our understanding of the rise of early animals.
A distinctive biomarker and organic-walled microfossils assemblage from oil shales in a terminal Neoproterozoic-lower Cambrian evaporite succession: evidence from the Salt Range Formation in East Salt Range, northern PakistanTOP
F.W. Meng1, P. Ni2, N. Ahsan3, S. Ur Rehman4, W.H. Liu5
1State Key Laboratory of Palaeobiology and Stratigraphy, Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences, Nanjing, China. Email: fwmeng@nigpas.ac.cn
2School of Earth Sciences and Engineering, Nanjing University, State Key Laboratory for Mineral Deposits Research, Institute of Geo-Fluid Research, Nanjing, 210093, China.
3Institute of Geology, University of the Punjab, Lahore, Pakistan.
4Department of Earth Sciences, University of Sargodha, Sargodha, Pakistan.
5Institute of Geological Sciences, Polish Academy of Sciences, Warsaw, Poland.
A study of the terminal Neoproterozoic-early Cambrian palaeobiology in the India Block is focused on the molecular fossil record from oil shales in Salt Range, Pujab Basin, northern Pakistan. Biomarkers from oil shales have been extracted at the top of the Salt Range Formation, East Salt Range, Pujab Basin, northern Pakistan, and analyzed using gas chromatography-mass spectrometry. The biomarker assemblage indicates distinctive and significant algae and bacteria contribution similar to other evaporitic basins during the time. Dominance of C29 sterane signifies that green algae were important primary producers, and similar to the Precambrian Huqf Group oils and solid bitumens from the Sichuan Basin, South China. The community of algae and other taxa, thrived in a thin oxygenated layer in the upper water column. However, the presence of an underlying euxinic water column is demonstrated by the presence of gammacerane. Excellent preservation of the biomarkers and organic-walled microfossils is indicated by sterane and triterpane molecular ratios with preferences for biological stereoisomers. The entire biomarker assemblage is indicative of a restricted, salinity-stratified depositional environment, possibly including the presence of extensive, anoxygenic microbial mats. Some of the observed characteristics mirror those of other coeval successions in Oman, India and South China, but the particular combination of features appears to be unique to this setting.
The relationship between reproductive mode and intra-specific competition in Ediacaran Avalonian macrofossil assemblagesTOP
E.G. Mitchell1, A.G. Liu2
1Department of Zoology, University of Cambridge, UK. Email: emily.g.h.mitchell@gmail.com
2Department of Earth Sciences, University of Cambridge, UK.
Reproductive traits are key to an organism’s capacity to adapt and evolve, but little is known about the influence of reproductive modes on selection pressures within the Ediacaran macrobiota. Previous analyses of ~580-560 Ma Avalonian palaeocommunities from Newfoundland (Canada) and Charnwood Forest (UK) found that inter-specific resource competition was rare, and weak where present. Independent spatial analyses predicted stoloniferous reproduction within some Avalonian rangeomorph populations, which has seemingly been corroborated by fossil evidence suggesting that filamentous stolons facilitate persistent connectivity even between large specimens. These latter results offer a possible explanation for the observed low resource competition, whereby physical connection of proportions of Avalonian populations via stolons enabled resource sharing, thus reducing intra-specific competition. In this study we test this hypothesis by investigating the relationship between reproductive mode and intra-specific competition for 12 Avalonian palaeocommunities.
Spatial point process analyses (SPPA) were used to identify taxon dispersal patterns within palaeocommunities, with analyses of population size-distributions used to constrain methods of reproductive behaviour. Assessment of whether taxa utilised waterborne and/or tethered/stoloniferous reproduction was performed by analysis of the directionality of dispersal clusters using K-measures, calibrated against models of water-dispersal mechanisms in extant taxa. We independently assessed our inferred reproductive behaviours using the Coefficient of Dispersal to identify significant intra-specific morphological variation, which can distinguish between asexual and sexual reproductive strategies. Together these methods enabled reconstruction of the reproductive modes of 13 taxa. The presence and strength of intra-specific competition within the communities was determined using SPPA, which found only rare instances of such competition. Carrying capacity was rarely reached in the studied populations, suggesting reduced selection pressures within these Avalonian communities. Combining these analyses enables us to use General Linear Model regressions to investigate the extent to which asexual reproduction influenced intra-specific competition within Avalonian communities.
Algal cysts - not animal embryos - are recognized among Ediacaran microfossils from the Doushantuo Formation, South ChinaTOP
M. Moczydłowska1, P. Liu2
1Uppsala University, Department of Earth Sciences, Palaeobiology, Villavägen 16, SE 752 36 Uppsala, Sweden. Email: malgo.vidal@pal.uu.se
2Institute of Geology, Chinese Academy of Geological Science, Beijing 100037, China.
Organic-walled microfossils from the early-middle Ediacaran Doushantuo Formation studied in several sections in the Yangtze Gorges area, South China, display lavishly ornamented cyst-like vesicles of high taxonomic diversity. Microfossils are diagenetically silicified in chert nodules and were studied in thin sections. Seven species of Appendisphaera, Mengeosphaera, Tanarium, Urasphaera, and Tianzhushania, among many other recorded taxa, preserved internal bodies (single spheroidal structure) and dividing cells (multiple, undifferentiated spheroids) inside the vesicles, which are reproductive cyst stages. Various ontogenetic developmental phases are observed in the same taxa, having a few to hundreds dividing cells within the same morphologically and dimensionally cyst. We interpret the new evidence as indicating the algal affinities of studied taxa and question the previously suggested interpretation of being animal cyst and embryos for Tianzhushania and Appendisphaera. Appendisphaera has been originally considered to be algal cyst by various lines of evidence and new specimens preserved in the late reproductive stage confirm this affinity. The first record of late developmental stage of completely preserved specimen of Tianzhushania demonstrates the interior of cyst containing identical cells cluster and without any cavity that is diagnostic for recognizing algal vs animal cysts. Our inferences on biological affinities of microfossils are based on morphology, reproductive characters, topology of cells, and biochemical properties of refractory biopolymers which comprised the vesicle wall. Collectively, all features are characteristic of algal clades by comparison with extant biota. The morphologic complexity of microfossils and many individual features, alongside their reproductive stages are in concert with algal morphotypes and representing classes known to exist at the time. Coeval animal grades are only represented by sponge, cnidarians, and placozoans, which have never produced the cysts of similar complexity, neither the holozoans that were also suggested as possible organisms forming such fossilized cysts.
The new acritarch record of the Ediacaran Bocaina Formation, Corumbá Group, BrazilTOP
L. Morais1,5, T. Fairchild2, J.M. Leme2, I.D. Rudnitzki3, B.T. Freitas4, P.C. Boggiani2, R.I.F. Trindade5
1Postdoctoral Researcher, Geochemistry and Geotectonics Program, Institute of Geosciences, Universidade de São Paulo - USP, Rua do Lago, 562, Cidade Universitaria, CEP: 05508-080, São Paulo, Brazil. Email: lumoraiss@ymail.com
2Institute of Geosciences, University of São Paulo, São Paulo, Rua do Lago, 562, Cidade Universitária, CEP 05508-080, Brazil.
3Department of Geology, Universidade Federal de Ouro Preto - DEGEO/UFOP. Morro do Cruzeiro, s/n Bauxita, CEP: 35400-000, Ouro Preto, Brazil.
4Geology Laboratory, School of Technology, Universidade de Campinas - UNICAMP, Rua Paschoal Marmo, 1888, Jardim Nova Italia, CEP: 13484-332, Limeira, Brazil.
5Institute of Astronomy, Geophysics and Atmospheric sciences. University of São Paulo, Rua do Matão, 1226, São Paulo, SP CEP: 05508-090, Brazil.
The Bocaina Formation crops out in the state of Mato Grosso do Sul, SW Brazil, beneath the black limestones and shales of the Tamengo Formation, where the important Corumbella and Cloudina are found (> 542 Ma). Zircons from interbedded tuffs from the upper part of the Bocaina Formation were dated at 555.18 ± 0.30 Ma (ID-TIMS U-Pb). Both formations are part of the Ediacaran Corumbá Group. The unit in question is essentially dolostone deposited in an evaporitic lagoonal context, with common stromatolites and, in the upper part, phosphorite intercalations that yielded the microfossils described here. The assemblage of microfossils includes sphaeromorphs, netromorphs, and acanthomorphs acritarchs, in addition to two new categories of organic microfossils, possibly representing a basal radiolarian and a multicellular eukaryote, respectively. Presently known acritarchs include members of genera that have proved useful in Neoproterozoic biostratigraphy, such as Tanarium (600-535 Ma) and Trachyhystrichosphaera (1030-600 Ma), reported here for the first time in the Corumbá Group. Of the new categories, one comprises two specimens made up of four organic rings that form a radially symmetric globular framework (less than 50 μm in diameter), reminiscent of structures found in radiolarians. To date, the oldest known radiolarians occur in earliest Cambrian black cherts of the Yangtze Platform, China. If the affinity with radiolarians can be fully corroborated, this would mean that a heterotrophic planktonic protistan lifestyle can be extended back into the Ediacaran. The other novel category consists of two larger (c. 90 μm in diameter), more complex specimens, similar to the acanthomorphic acritarch Eotylotopalla delicata, but differs from this taxon in having two distinct openings. This feature is here interpreted as indicative of a multicellular affinity, possibly a stage in the life cycle of an early multicellular organism (egg, embryo, juvenile or even adult). The diversity of microfossils found in the Bocaina Formation promises to help resolve its biostratigraphic position with respect to other important Ediacaran fossiliferous units and bolsters its paleobiological importance on a global scale.
Funding Project: FAPESP (2016/06114-6); (2017/22099-0)
What is the age of the earliest Cambrian polycystine radiolarians and what do they look like? Insights from the Gorny Altai (South of western Siberia)TOP
O.T. Obut1,2, T. Danelian3
1Trofimuk Institute of Petroleum Geology and Geophysics SB RAS, akad. Koptyug av., 3, Novosibirsk, Russia. Email: olgaobut@gmail.com
2Novosibirsk State University, Piragova st., 2, Novosibirsk, Russia.
3Université Lille, CNRS, UMR 9198 - Évo-Éco-Paléo, F-59000 Lille, France.
Molecular phylogenies suggest that polycystine radiolarians (the ones secreting aesthetically pleasant and delicate siliceous skeletons) are one of the oldest lineages amongst the Rhizarian supergroup of unicellular heterotrophic eukaryotes (Sierra et al., 2013). Precise knowledge of their oldest fossil record is crucial, not only for the calibration of molecular trees, but also for improved understanding of their involvement in the evolution of food webs during the Ediacaran-Cambrian transitional interval, as well as for the initiation of siliceous biomineralization in eukaryotic lineages (Danelian & Morreira, 2004).
The presence of polycystine radiolarians in the Middle Cambrian was firmly established 20 years ago, based on a well-preserved and diverse microfauna extracted from Templetonian - Undillan strata of Australia (Won & Below, 1999). This fauna is represented by four distinct clades, three of which are part of the primitive radiolarian order Archaeospicularia (Dumitrica et al., 2000); this concerns the families Archeoentactiniidae and Palaeospiculidae, as well as the echidninid genus Parechidnina. The fourth clade is represented by the genus Protobiramus which is currently regarded as incertae sedis at both the order and family levels (Caridroit et al., 2017).
However, the record of early Cambrian radiolaria and their morphological and anatomical appearance is a matter of ongoing controversy (see Aitchison et al., 2017). Indeed, as it was argued convincingly by Maletz (2011, 2015, 2017) the scarce specimens presented by Braun et al. (2007) and more recently by Cao et al. (2014) as radiolarians yielded from lower Cambrian levels of South China are likely to be contamination from younger material.
Nevertheless, three recent papers presenting radiolarians from Lower Cambrian strata of South China have revived the debate by questioning the archaeospicularian-type skeleton structure for the oldest polycystine radiolarians (Zhang & Feng, 2019; Chang et al., 2019) and by claiming, on the contrary, a Spumellarian origin (Ma et al., 2019).
Cambrian radiolarians extracted from the Shashkunar pelagic sedimentary sequence that crops out at the Ak-Kaya section of the Gorny Altai (southern part of western Siberia) provides useful insights to this debate. The presence of polycystine radiolarians was initially reported nearly 20 years ago (Obut & Iwata, 2000; Zybin et al., 2000); these were extracted from siliceous mudstones with the use of dilute hydrofluoric acid. However, their preservation was rather poor and did not allow the observation of any internal structures. They are now considered as being part of the Archaeospicularia (Noble et al., 2017).
A Russian-French collaborative project, initiated in 2010, allowed re-examination of the radiolarian-bearing strata cropping out at the Ak-Kaya section. Results established the presence of archeontactiniid radiolarians in an 8 m-thick interval of bedded cherts situated at the upper part of the Shashkunar Formation (Pouille et al., 2011). Investigations during this project allowed also the discovery of lower Botoman trilobites from two carbonate horizons situated a few tens of meters below the radiolarian-bearing chert interval (Korovnikov et al., 2013; Sennikov et al., 2017).
We have recently obtained additional material from the siliceous mudstone beds situated in the upper siliceous interval of the Ak-Kaya section. Although the preservation of the extracted radiolarians is average to poor, it is possible to observe spherical forms devoid of any internal spicule or outer spines, made of a fairy thick shell wall composed of irregularly interwoven threadlike elements. Some of the specimens recall the echidninid genus Parechidnina.
The novelty of our results consist in the fact that extensive processing of the trilobite-bearing limestones with dilute acetic acid yielded several tens of fairly well-preserved radiolarian specimens, as well as diverse forms of siliceous sponge spicules and the SSF species Rhombocorniculum cancellatum. The latter is known worldwide from the late Atdabanian - early Botoman interval and is compatible with the early Botoman age provided by trilobites. The radiolarians extracted from these limestones display clearly all characters of the archaeospicularian genus Archeoentactinia; they are characterized by a 6-rayed point-centered internal spicule and a spherical to subspherical shell constructed by an irregularly interwoven meshwork. The shell is made of thin, thread-like, delicate elements placed at various angles in such a way to form a complex three-dimensional meshwork. The eccentric position of the point-centered spicule, somehow tangential to the shell wall, is clearly visible on some specimens. A second morphotype that may also be assigned to genus Archeoentactinia displays numerous arch-shaped elegant bars constructing an irregularly interwoven meshwork and forming an imperfectly spherical shell. The thin rays of the internal spicule protrude through the shell as long outer spines.
In summary, all radiolarians extracted from both the upper siliceous interval and the trilobite-bearing limestones belong respectively to the echidninid and archeoentactinian lineages of Archaeospicularia. The absence of any palaeospiculids in our material is noteworthy. They are early Botoman in age (early Cambrian Stage 4) and they represent the oldest radiolarians known so far. Our results confirm the hypothesis that the earliest polycystine radiolarians were characterized by spicular skeletons and archaeospicularian type of body plans and that they built their shells by a three-dimensionally interwoven meshwork of spines and bars.
O. Obut’s research was supported by the Ministry of Education and Science of the Russian Federation grant n° 14.Y26.31.0018. T. Danelian thanks the Région Hauts-de-France and the Ministère de l’Enseignement Supérieur et de la Recherche (CPER Climibio), and the European Fund for Regional Economic Development for their financial support.
Hydrothermal influence on barite precipitates in the basal Ediacaran Sete Lagoas cap carbonate, São Francisco Craton, central BrazilTOP
J. Okubo 1,3, Y. Igorevich Klyukin 2,3, L. Veríssimo Warren1, R.J. Bodnar3, B.C. Gill3, Shuhai Xiao 3
1Department of Applied Geology, Institute of Geosciences and Exact Sciences, São Paulo State University (Unesp), Rio Claro-SP, Brazil. Email: juliana.okubo@unesp.br
2Department of Earth & Atmospheric Sciences, University of Alberta, Edmonton, Canada.
3Department of Geosciences, Virginia Tech, Blacksburg, Virginia 24061, USA.
Most Precambrian sediment-hosted barites are usually formed by diagenetic, hydrothermal exhalation, or methane-seepage processes. However, seafloor barite precipitates and void-filling barite cements in basal Ediacaran cap carbonates have been interpreted as sedimentary and early diagenetic in origin. Barite crystals that precipitated syndepositionally or authigenically from porewaters in direct communication with seawater can potentially offer insights into Neoproterozoic ocean geochemistry in the aftermath of the terminal Cryogenian (or Marinoan) Snowball Earth glaciation.
In this study, we investigated void-filling barite cements in the basal Ediacaran cap dolostone of the Sete Lagoas Formation, Bambuí Group, central Brazil. Our goal was to investigate the origin and diagenetic history of the Sete Lagoas barite by addressing the following questions: (i) whether the barite precipitated from porewaters in direct connection with seawater and hence sulfate was ultimately sourced from seawater, (ii) whether barite cements experienced any subsequent hydrothermal alteration, and (iii) if hydrothermal alteration did occur, what is the nature of the hydrothermal fluids.
Sete Lagoas barite occurs as veins and as major void-filling cement in multiple horizons (Fig. 10A), sometimes interbedded with carbonate fans. Barite is present as radiating bladed crystals forming rosettes which grow inward from both the top and bottom of the vug (Fig. 10B-D), as isolated crystals in the matrix, and as inclusions within carbonate fan crystals and minor void-filling cements, sometimes partially replacing carbonate fans.
|
 Figure 10.—Mesoscopic and microscopic features of a barite layer. A. Void-filling barite in the outcrop, commonly forming rosettes, growing inward from both the top and bottom of the vug. B. Dolomitic matrix (top) and radiating bladed barite crystals (bottom). C. Detail of B showing radiating bladed crystals of barite. Plane polarized light microscopy (PPL). D. Enlarged view of (C), showing trails of intergranular and intragranular secondary (?) fluid inclusions perpendicular to the growth direction of barite crystals. PPL. Figure 10.—Mesoscopic and microscopic features of a barite layer. A. Void-filling barite in the outcrop, commonly forming rosettes, growing inward from both the top and bottom of the vug. B. Dolomitic matrix (top) and radiating bladed barite crystals (bottom). C. Detail of B showing radiating bladed crystals of barite. Plane polarized light microscopy (PPL). D. Enlarged view of (C), showing trails of intergranular and intragranular secondary (?) fluid inclusions perpendicular to the growth direction of barite crystals. PPL.
|
|
Sulfur isotope compositions of coexisting barite and carbonate-associated sulfate or CAS (see location of the isotope analyses in Fig. 11) show similar values, suggesting a similar source, perhaps both derived from seawater sulfate. Assuming that CAS was sourced from seawater sulfate, we infer that the Sete Lagoas barite was precipitated from porewaters in connection with seawater and hence the source of sulfate ultimately came from the seawater.
|
 Figure 11.—X-ray CT three-dimensional reconstruction of a Sete Lagoas sample that contains a layer of void-filling barite crystals (bottom half of the specimen) and a layer of the host rock (top half of the specimen). Barite is segmented as green color, micritic matrix in the barite layer as pink color, and laminated dolostone in the host rock as blue color. A-B and C-D are two different views of the same specimen, with A and D showing only the barite component of the specimen. δ34SCAS and δ34Sbarite values are marked on the specimen. The specimen is oriented in stratigraphic up direction. Figure 11.—X-ray CT three-dimensional reconstruction of a Sete Lagoas sample that contains a layer of void-filling barite crystals (bottom half of the specimen) and a layer of the host rock (top half of the specimen). Barite is segmented as green color, micritic matrix in the barite layer as pink color, and laminated dolostone in the host rock as blue color. A-B and C-D are two different views of the same specimen, with A and D showing only the barite component of the specimen. δ34SCAS and δ34Sbarite values are marked on the specimen. The specimen is oriented in stratigraphic up direction.
|
|
Carbonaceous material (CM) in barite crystals and CM in fluid inclusions trapped in barite crystals share a similar thermal history, with a peak temperature in the range of 206-257°C. CM was not detected in the host rock, possibly due to pervasive dolomitization, thermodegradation or the fine-grained nature of the host rock. Temperatures and salinity, evaluated by fluid inclusion study, are both higher than seawater. We suggest that hydrothermal activity may have modified the original composition of the Sete Lagoas barite. This activity may be linked to MVT mineralization in the study area. Thus, the Sete Lagoas barite may have been subsequently modified by hydrothermal activities, after its precipitation from seawater or porewaters in communication with seawater. From this perspective, a thorough diagenetic assessment must be done before the Sete Lagoas barite can be used to infer seawater geochemistry and environment in the aftermath of Marinoan Snowball Earth.
This research was sponsored by FAPESP (grant 2015/07391-0 and 2017/19023-1) and NASA Exobiology and Evolutionary Biology Program (80NSSC18K1086).
The timescale of animal evolutionTOP
J.E. O’Reilly1, D. Pisani1, P.C.J. Donoghue1
1School of Earth Sciences, University of Bristol, Life Sciences Building, Tyndall Avenue, Bristol BS8 1TQ, UK. Email: philip.donoghue@bristol.ac.uk
The timing of origin and early evolution of animals is bound up with whether we interpret the Cambrian Explosion to reflect an explosive diversification of animals or an explosive increase in fossils, relative to the foregoing Neoproterozoic. There can be no doubt that animal phyla diverged prior to their appearance in the stratigraphic record but the same palaeontological and phylogenetic evidence underpins competing views that lie at and between the extremes of crown animals emerging in the deep Neoproterozoic versus limited entirely to the Phanerozoic. However, fossil evidence alone cannot establish a timescale of animal evolution, the only viable methodology for which remains the molecular clock. It is perhaps surprising that most scenarios for the emergence of animal diversity remain calibrated to the fossil record but this likely reflects that lack of confidence which stems from the implied extent of early animal evolution unrepresented by the fossil record – but also their lack of consistency, with some studies differing by as much as hundreds of millions of years. This lack of consistency between molecular clock studies is not surprising since divergence time methods are now diverse and use palaeontological data in different ways. Here we attempt to benchmark molecular clock methods using a common suite of molecular and morphological data, living and fossil taxa, and phylogenetic hypotheses. In comparison to conventional node calibration, we have undertaken a morphological clock analysis including both living and fossil taxa, a combined molecular and morphological Total Evidence Dating (TED) analysis, and an analysis calibrated using the Fossilised Birth-Death (FBD) integrated diversification and fossil sampling model. In particular, it has been anticipated that methods which incorporate more palaeontological data and are guided by the stratigraphic distribution of included fossil species, such as TED and FBD, might result in evolutionary timescales that are in closer agreement with the fossil record. However, in their application to the timing of origin of animals, this expectation is not met. While the results from application of these and other methods differ in their precision, they are agreed that animals diverged long before their appearance in the fossil record.
Early evolution of eukaryotes after the Marinoan glaciation: data from the Ediacaran Doushantuo Formation at the Jiulongwan section, Yangtze Gorges area, South ChinaTOP
Q. Ouyang1, C. Zhou1, C. Guan1
1CAS Key Laboratory of Economic Stratigraphy and Palaeogeography, Nanjing Institute of Geology and Palaeontology, and Center for Excellence in Life and Palaeoenvironment, Chinese Academy of Sciences, Nanjing 210008, China. Email: qouyang@nigpas.ac.cn
The Ediacaran Period witnessed the early diversifications of marine eukaryotes after the terminal Cryogenian global glaciation. These stepwise evolutionary events include a rapid radiation of microscopic acritarchs and multicellular algae in the early-mid Ediacaran, the succeeding diversification of macroscopic Ediacaran biota in the mid-late Ediacaran, and the appearance of biomineralized skeletal organisms in the Ediacaran-Cambrian transition interval (Narbonne et al., 2012). Palaeontological investigations during the past decades of years have also demonstrated the coexistences of microscopic acanthomorphic acritarchs with the megascopic Ediacara-type organisms (e.g., Golubkova et al., 2015; Ouyang et al., 2017), and the late Ediacaran tubular fossils like Cloudina with the early Cambrian small shelly fossils (e.g., Zhu et al., 2017; Cai et al., 2019).
But questions concerning when and how the first Ediacaran eukaryotes began to thrive and dominate the marine ecosystem still remain. Acanthomorphic acritarchs have been recovered from around the world, including Ediacaran successions in southern Norway, Svalbard, Siberia, South China, central and southern Australia, Lesser Himalaya, East European Platform, Vindhyan Basin, and northern Mongolia (e.g., Spjeldnaes, 1963; Knoll, 1992; Zhang et al., 1998; Grey, 2005; Vorob’eva et al., 2009; Golubkova et al., 2010; Joshi & Tiwari, 2016; Prasad & Asher, 2016; Anderson et al., 2017). However, the first appearance datum (FAD) of acanthomorphic acritarchs varies among different sections. In Australia, Siberia and East European Platform, the FAD of the organic-walled acritarchs are hundreds or even thousands of meters above the base of the Ediacaran System (Grey, 2005; Moczydłowska & Nagovitsin, 2012). In South China, however, the FAD of the permineralized acritarchs is much lower, i.e. ~ 10 meters above the Ediacaran basal boundary (Zhou et al., 2007). Even if taken into consideration that the lower Ediacaran Doushantuo Formation in South China is relatively condensed, the FAD of acanthomorphic acritarchs in South China is still much lower than those in other localities. Here we present our recent discoveries of microscopic eukaryotes from the Doushantuo Formation in the Yangtze Gorges area, South China, and discuss briefly on their stratigraphic range, correlation, and evolutionary pattern.
Our palaeontological investigation shows that at the Jiulongwan section in the Yangtze Gorges area, South China, the lowest occurrence horizon of the Ediacaran eukaryotic microfossils is only 2.8 m above the top of cap dolostone (Zhou et al., 2019). This is the lowest horizon yielding eukaryotic microfossils in the Ediacaran strata ever reported in South China. Microfossils from this horizon include acanthomorphic acritarch Tianzhushania, which is the most common taxon in South China, and several multicellular algae species such as Wengania minuta and Sarcinophycus radiatus, as well as filamentous cyanobacteria. These fossils range upward through the lower part of the Doushantuo Formation at the Jiulongwan section, with the diversity of acanthomorphic acritarchs increases upsection. Within ten meters above the top of cap dolostone, more than ten genera of acanthomorphic acritarchs appear, including the widely distributed Appendisphaera, Knollisphaeridium, and Weissiella, and domestic taxa such as Dicrospinasphaera and Yinitianzhushania. The present data also show that half of the acanthomorphic acritarch taxa at the Jiulongwan section appear within 15 m above the top of cap dolostone.
These findings are consistent with the results of a very comprehensive palaeontological study of the Doushantuo acanthomorphic acritarchs recently published by Liu & Moczydłowska (2019). Although no detailed stratigraphic height is given, their data show that the FAD of acanthomorphic acritarchs is less than ten meters above the basal boundary of the Ediacaran System at the Chenjiayuanzi section. The Chenjiayuanzi section is only several kilometers away from the Jiulongwan section, and the lower part of the Doushantuo Formation at these two sections are completely correlatable (Liu et al., 2014). Taxa appeared at the lowest horizon at the Chenjiayuanzi section are Knollisphaeridium, Tianzhushania, and Yinitianzhushania (Liu & Moczydłowska, 2019), which are comparable with the microfossil assemblage discovered from approximately the same horizon at the Jiulongwan section.
The present available microfossil data indicate that eukaryotic life recovered and flourished soon after the termination of the Marinoan glaciation. In the Yangtze Gorges area, two zircon U-Pb ages of 635.2 ± 0.6 Ma and 632.5 ± 0.5 Ma have been reported for ash beds of 2.3 m and 9.5 m above the base of the Doushantuo Formation, respectively (Condon et al., 2005), indicating that within millions of years a new marine ecosystem dominated by microscopic eukaryotes has been established. This marine ecosystem became more complex with the later diversification of microscopic organisms (e.g., Xiao et al., 2014; Liu & Moczydłowska, 2019) and appearance of macroscopic and morphologically differentiated eukaryotes in local areas (Yuan et al., 2011) during early-mid Ediacaran.
It is worth noting that the occurrence of eukaryotic fossils above the glacial deposits is very likely to be controlled by particular taphonomic window. Microfossils from the Doushantuo Formation are all permineralized, and most of them are found from chert nodules or phosphorites. Since acanthomorphic acritarchs were discovered from the lowest chert nodule horizon at the Jiulongwan section, the possibility that the absence of microfossils below this horizon is due to the lack of suitable preservational condition cannot be completely excluded.
The FAD of acanthomorphic acritarchs has already been suggested as a potential criterion for the stage-level subdivision of the Ediacaran System for it represents a significant evolutionary event (e.g., Liu et al., 2014; Xiao et al., 2016). However, as the first appeared acanthomorphic species, Tianzhushania spinosa has very restricted spatial distribution. Therefore, integrated biostratigraphic and chemostratigraphic approaches, as well as event-stratigraphic marker need to be considered for the subdivision and correlation of the lower Ediacaran System.
This study was supported by the Strategic Priority Research Program (B) of Chinese Academy of Sciences (XDB18000000), National Key R & D Program of China (2017YFC0603101), and National Natural Science Foundation of China (41672027).
Current status of the Ediacaran-Cambrian Bilara Group, Marwar Supergroup, IndiaTOP
S.K. Pandey1, N. Bykova2, M. Sharma1, G.A. Karlova2, A.H. Ansari1, Y. Kumar1, S. Ahmad1, M.K. Pandit3
1Birbal Sahni Institute of Palaeosciences, 53 University Road, Lucknow, India. Email: sangeology@yahoo.co.in, skpandey@bsip.res.in
2Trofimuk Institute of Petroleum Geology and Geophysics, Siberian Branch Russian Academy of Sciences, Prospekt Akademika Koptyuga 3, Novosibirsk, Russia.
3Department of Geology, University of Rajasthan, Jaipur India.
On the edge of the Neoproterozoic Era, the Earth experienced diverse and consequential changes in the lithosphere, biosphere, atmosphere, and hydrosphere. It is marked by major carbon cycle perturbations, transient oxygenation of the deep ocean, and transformative biological innovations (Erwin et al., 2011, Lenton et al., 2014). Life has got big after the Marinoan and Gaskiers glaciations in the Ediacaran Period and more complex and diverse after the Cambrian Explosion (Marshall, 2006). Thus, the study of changes across the Precambrian-Cambrian boundary is a significant move to encounter above mentioned changes in the Earth’s history. In order to achieve that goal, it is essential to identify the successions preserving this crucial time of the Earth’s history. The present study is focused on establishing a new horizon of the Ediacaran-Cambrian succession in peninsular craton of India i.e. the Bilara Group of the Marwar Supergroup on the basis of new findings. For a long time, the Krol and Tal formations of the Lesser Himalaya used to be known as the only well established Precambrian- Cambrian succession in India (Aharon et al., 1987; Bhatt & Mathur, 1990; Hughes, 2016). The Bilara Group is calcareous unit, which sandwitched between underlying siliciclastic Ediacaran Jodhpur Group and overlying Cambrian Nagaur Group (Pareek, 1981, 1984). The Bilara Group of rocks mainly composed of limestone, dolostone intercalated with claystone/siltstone or chert bands. Previous stable isotope studies suggest the Precambrian-Cambrian boundary lie within the Bilara Group (Maheshwari et al., 2003). Recently, robust C and O stratigraphy reveled that the Bilara carbonate witnessed with high amplitude negative carbon excursion (Shuram Excusrsion) (Ansari et al., 2018). About 23 m thick Barna mine section, includes the upper part of the Gotan Limestone, which unconformably overlain by the Pondlo Dolomite. The siltstone horizon is dated by the U-Pb method, which provides 544 Ma age (maximum depositional age) (Wang et al., 2019). Moving towards the top of the mine section, about 5 meters above the dated horizon, C and O stratigraphy suggest the probable signature of the Precambrian-Cambrian boundary. Ediacaran fossils and Cambrian trace fossils have already been reported from the underlying Sonia Sandstone and overlying Nagaur Sandstone. Therefore, in the current scenario, the Bilara Group includes Late Ediacaran and Lower Cambrian rock succession.
Here we demonstrate the new occurrence of the Cambrian-type skeletal fossil Cambrotubulus sp. from the Upper Gotan Limestone of the Barna-I Mine. So far, two fragmented specimens have been recovered from the dissolution of carbonate rocks from the layer which lies about 8 meters from the base of the mine section, and two meters below of the U-Pb age of 544 Ma. The fossils of Cambotubulus are associated with the terminal Ediacaran-early Cambrian time (Cai et al., 2019: Rogov et al., 2015; Zhu et al., 2017). Thus, the presence of Cambrotubulus in the Bilara Group of India places a hard constraint on the age of the Gotan Limestone, as well as opens the new opportunities for the search of the Ediacaran and Cambrian-type fossils in the Ediacaran successions of the Bilara Group. Overall, integrated palaeontological, chemostratigraphic and geochronological investigations suggested that the Bilara Group is witnessing the Ediacaran-Cambrian transformation of the Earth’s ecosystem, which needs thorough attention in the future, for establishing the timeline of these events in the Bilara Group.
Organic-walled microfossils from the Lower Neoproteorozoic Tongjiazhuang Formation in western Shandong, North China and their biostratigraphic significanceTOP
K. Pang1, G. Li2, L. Chen2
1State Key Laboratory of Palaeobiology and Stratigraphy, Nanjing Institute of Geology and Palaeontology and Center for Excellence in Life and Paleoenvironment, Chinese Academy of Sciences, Nanjing 210008, China. Email: kepang@nigpas.ac.cn
2College of Earth Science and Engineering, Shandong University of Science and Technology, Qingdao 266590, China. Email: leichen@nigpas.ac.cn
Neoproterozoic successions in western Shandong of North China contain fine-grained siliciclastic rocks which have the potential to preserve organic-walled microfossils. Unfortunately, their abundance and diversity of organic-walled microfossils have long been overlooked and the exact depositional age of these units has not been well constrained. In this study, we have systematically investigated organic-walled microfossils, using low-manipulation maceration techniques, from the Tongjiazhuang Formation, Tumen Group in western Shandon, North China. A well-preserved and diversified microfossil assemblage, characterized by abundant smooth-walled sphaeromorphic acritarchs and cyanobacterium-like filamentous forms and relatively low abundance of more complex acritarchs, is obtained from the Tongjiazhuang Formation. A total of 40 microfossil taxa belonging to 20 morphological genera have been identified, most of which are described for the first time from the Tongjiazhuang Formation of the Tumen Group in western Shandong, including Simia sp., Trachyhystrichosphaera aimika, T. botula, Germinosphaera bispinosa, unnamed sp. A, Jacutianema solubila and Pololeptus rugosus. The Tongjiazhuang assemblage revealed in this study is much more diverse than previously reported and well comparable to the Tonian Liulaobei and Gouhou assemblages in the Xuhuai region. Particularly, the occurrence of T. aimika, T. botula, G. bispinosa, J. solubila and P. rugosus indicates a late Mesoproterozoic to Tonian age for the Tongjiazhuang Formation. In addition, the occurrences of macroscopic carbonaceous compressions, including a Chuaria-Tawuia assemblage from the Tongjiazhuang Formation and Tawuia and Sinosabellidites from the Shiwangzhuang Formation, are consistent with geochronological data (youngest detrital zircon ages of 1063 Ma and a Rb-Sr whole rock isochron age of 910.2 Ma), suggesting a Tonian age for the Tongjiazhuang, Fulaishan, and Shiwangzhuang formations of the middle and upper Tumen Group, while the Heishanguan and Erqingshan formations of the lower Tumen Group are likely late Mesoproterozoic to Tonian in age. Thus, available biostratigraphic data, lithostratigraphic correlation, and geochronological data suggest that the whole Tumen Group is late Mesoproterozoic to Tonian in age, rather than Cryogenian or Ediacaran. Our study confirms the existence of the ‘Great Unconformity’ in western Shandong, between the upper Mesoprotoerozoic to Tonian Tumen Group (represented by the Tongjiazhuang Formation at the Tangtou section or the Shiwangzhuang Formation in some other areas of western Shandong) and the lower Cambrian Liguan Formation. This study significantly improves our knowledge about the diversity of the Tonian biosphere.
Cloudinids surviving the Ediacaran-Cambrian transitionTOP
T.Y. Park1, J. Jung1, M. Lee1, S. Lee2, Y.Y. Zhen3, N.C. Hughes4
1Division of Earth-System Sciences, Korea Polar Research Institute, 26 Songdomirae-ro, Yeonsu-gu, Incheon 21990, Korea Republic. Email: typark@kopri.re.kr
2School of Earth, Atmospheric and Life Sciences, Univeristy of Wollongong, Northfields Ave, Wollongong, NSW 2522, Australia.
3Geological Survey of New South Wales, W.B. Clarke Geoscience Centre, 947-953 Londonderry Road, Londonderry, NSW 2753, Australia.
4Department of Earth Sciences, University of California, Riverside, CA 92521, USA.
Cloudina is an iconic fossil of the terminal Ediacaran skeletal fauna, but its phylogenetic affinity remains unclear. Cloudina is known to have demised at the Ediacaran/Cambrian boundary, and thus often considered as an index taxon of the terminal Ediacaran. Cambroctoconus is an octoradial cup-shaped animal occurring from the Cambrian Series 2 to Miaolingian of Greenland, Kyrgysztan, China and Korea. It was originally reported as a stem-group cnidarian, but a possibility of this animal being related to crown-group octocorals has been suggested. A recent study revealed that Lipopora and Tretocylichne from the Cambrian Stage 4 and Stage 5, respectively, of Australia are also closely related to Cambrocotocus, implying a remarkable radiation of this octoradial animal group during the Cambrian. A new observation using MicroCT shows that Lipopora has a funnel-in-funnel shaped morphology which is reminiscent of Cloudina. Interestingly, a newly-recognized growth mode of Cambroctoconus also produced a funnel-in-funnel shape. This may indicate that Cloudina did survive the Ediacaran-Cambrian boundary, and gave rise to octoradial cup-shaped animals in the Cambrian.
Revisiting the Ediacaran-Cambrian boundary in the Ossa-Morena Zone (SW Iberia)TOP
M.F. Pereira1, I. Dias da Silva2, C. Gama1, J.B. Silva2
1Instituto de Ciências da Terra, Departamento de Geociências, Escola de Ciências e Tecnologia, Universidade de Évora, Portugal. Email: mpereira@uevora.pt
2Instituto D. Luiz, Departamento de Geologia, Faculdade de Ciências da Universidade de Lisboa, Portugal.
Some of the best outcrops of Iberia to study the Ediacaran-Cambrian boundary are located in the Ossa-Morena Zone. In the Crato-Campo Maior region (SW Iberia, Portugal), this stratigraphic boundary is marked by an angular unconformity (Gonçalves, 1971). The Ediacaran sedimentary rocks of the Série Negra Group (maximum depositional age of c. 545 Ma; Linnemann et al., 2008) are overlain by Early Cambrian strata. A folded foliation has been recognized in the Ediacaran metagreywackes, metapelites, black metachert, marbles and metabasic rocks (Pereira & Silva, 2002).
This deformation event is previous to the intrusion of c. 526-525 Ma granitic rocks (Barquete and Barreiros plutons; Pereira et al., 2011; Sánchez-García et al., 2013), and is not represented in the uncon-formable overlying Early Cam-brian strata including sandstone (maximum depositional age of c. 532 Ma; Pereira et al., 2011). At the base of the lower Cambrian stratigraphic section there is the Freixo-Segóvia volcanosedimen-tary complex consisting of felsic tuff interbedded with conglo-merate and rhyolitic-dacitic lava flow (Pereira et al., 2006). The conglomerate is composed of pebbles of volcanic rock (basalt, rhyolite, dacite and mafic and felsic tuff), granitic rocks, chert, quartzite, arkosic sandstone, greywacke and shale in a tuffaceous sandy matrix. This volcano-sedimentary complex is overlain by a sequence of sandstone and shale passing vertically to limestone beds which have been attributed to the lower Cambrian (Pereira et al., 2006) (see Fig. 12). An ongoing research project intends to date the volcanic rocks of the volcano-sedimentary complex using U-Pb zircon geochronology. The absolute dates determined from these volcanic rocks will provide the time framework for the calibration of the existing stratigraphic scheme based on regional correlation.
|
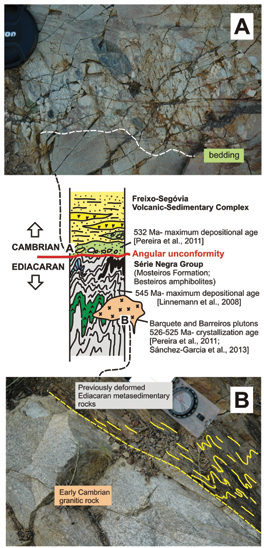 Figure 12.—Ediacaran-Cambrian stratigraphic section of the Ossa-Morena Zone (Crato-Campo Maior region, Portugal). A- Volcaniclastic conglo-merate is interbedded with sandy tuff beds (Early Cambrian Freixo-Segóvia Volcanic-Sedimentary Com-plex); B- Deformed metapelite (Edia-caran Série Negra Group) is cross-cut by a granitic rock (lower Cam-brian Barreiros pluton). Figure 12.—Ediacaran-Cambrian stratigraphic section of the Ossa-Morena Zone (Crato-Campo Maior region, Portugal). A- Volcaniclastic conglo-merate is interbedded with sandy tuff beds (Early Cambrian Freixo-Segóvia Volcanic-Sedimentary Com-plex); B- Deformed metapelite (Edia-caran Série Negra Group) is cross-cut by a granitic rock (lower Cam-brian Barreiros pluton).
|
|
This study is included in the research activity plan of the Instituto de Ciências da Terra (Universidade de Évora), Lithosphere Dynamics research group, with financial support from the Fundação para a Ciência e Tecnologia (Portugal), and of the project CGL2016-76438-P (Spanish MEIC).
Was the earliest deformation found in the Central-Iberian Zone Ediacaran Series solely caused by the Cadomian orogeny?TOP
M.F. Pereira1, C. Fernández2, A. Castro3, C. Rodríguez1,2
1Instituto de Ciências da Terra, Departamento de Geociências, Escola de Ciências e Tecnologia, Universidade de Évora, Portugal. Email: mpereira@uevora.pt
2Departamento de Ciencias de la Tierra, Universidad de Huelva, Spain.
3Instituto de Geociencias (IGEO), CSIC-UCM, Ciudad Universitaria, Madrid, Spain.
In the Central-Iberian Zone, the earliest deformation event found in the Ediacaran Series (i.e. Schist Greywacke Complex) is not represented in the unconformable overlying Early Ordovician strata. Traditionally there have been distinct views to explain the occurrence of this deformation, whose age is still uncertain. Some authors argue that the earliest deformation was caused by a Late Ediacaran contractional event (i.e. Cadomian; Talavera et al., 2012). The angular unconformity that separates the Lower and Upper Alcudian units has been regarded as evidence for a folding event without developing cleavage (Ortega Gironés & González Lodeiro, 1986; Palero, 1993). Other researchers believe that the earliest deformation was probably formed by rotational block faulting during a Cambro-Ordovician extensional event (Pereira et al., 2012). The abrupt changes in the thickness of the Ordovician basal units that overlain the Schist-Greywacke Complex are typical of deposition in extensional graben-horst systems (McDougall et al., 1987; Dias da Silva et al., 2015).
In the Cáceres region (Central Extremadura Batholith) the first cleavage observed in the Ediacaran Series has been attributed almost exclusively to Late Paleozoic (i.e. Variscan) deformation events which erased previous structures (Tena-Dávila Ruiz et al., 1980). However, towards the southeast, in the Miajadas-Navalvillar de Pela region, it has been described as the earliest deformation in the Ediacaran Lower Alcudian unit. Still, doubts persist about the age of the earliest deformation that for some authors corresponds to gently folding developed previously to the deposition of the Upper Alcudian unit (Matas et al., 1995) and for others, it represents an Ordovician deformation event (López Sopeña et al., 1991).
Recently, it was suggested that the earliest deformation found in the Ediacaran metasedimentary rocks of Cáceres mostly represents the response to an extensional deformation event that conditioned the emplacement at depth of the extensive cortege of coeval Ordovician plutonic bodies (Pereira et al., 2018). The new field, geochemical and geochronology findings make it possible to recognize the emplacement of intermediate magmas at shallow depths that gave rise to Ordovician extensive metamorphism due to heat transfer to the Ediacaran host rocks (Fig. 13).
|
 Figure 13.—Schematic reconstruction of the early-middle Ordovician plutonic-metamorphic belt of the Central Extremadura batholith (Cáceres), resulting in the development of: (A) a migmatitic foliation (Smig) in the Ediacaran Schist Greywacke Complex, which is associated with (B) tonalite layers with Ordovician crystallization age (modified from Pereira et al., 2018). Figure 13.—Schematic reconstruction of the early-middle Ordovician plutonic-metamorphic belt of the Central Extremadura batholith (Cáceres), resulting in the development of: (A) a migmatitic foliation (Smig) in the Ediacaran Schist Greywacke Complex, which is associated with (B) tonalite layers with Ordovician crystallization age (modified from Pereira et al., 2018).
|
|
Although both the field relations and the new radiometric ages are indicative of the existence of Ordovician deformation and metamorphism reaching high-grade conditions in the Central-Iberian Zone Ediacaran Series, we must not ultimately exclude the existence of previous structures, leaving some doubts to clarify in the scope of investigations still in progress.
This study is included in the research activity plan of the Instituto de Ciências da Terra (Universidade de Évora), Lithosphere Dynamics research group, with financial support from the Fundação para a Ciência e Tecnologia (Portugal), and of the project PGC2018-096534-B-I00 funded by Ministerio de Ciencia, Innovación y Universidades (Spain).
Late Proterozoic animals and the limits of fossil preservationTOP
L. Porras1, G. Wörheide1,2
1Department of Earth & Environmental Sciences & GeoBio-Center, Ludwig-Maximilians-Universität München, Germany. Email: L.Porras@lrz.uni-muenchen.de
2SNSB- Bayerische Staatssammlung für Paläontologie und Geologie, München, Germany.
The nature of the Cambrian explosion has been the subject of heated debate for decades. A popular interpretation poses that the sudden appearance is simply a consequence of the acquisition of hard parts such as jaw elements and thick cuticles during the diversification of bilaterian phyla. However, this may be a perception caused by our clearer understanding of bilaterian morphology and evolution compared to other non-bilaterian animals. A similar pattern of sudden appearance is also observed in the record of sponges; despite the lack of convincing Ediacaran spicules, Early Cambrian microfossil assemblages contain diverse spicule assortments, with even some types that are considered to be diagnostic of extant sponge orders. Furthermore, reports of Ediacaran ctenophores and cnidarians are still received with major skepticism by some authors. Here we explore character state distributions in non-bilaterian animals in an attempt to explain the conflict between molecular clocks and the scant record of early animals. We argue in favour of a taphonomic explanation imposed by early animal bodyplans. Most of the apomorphies accrued along the deepest splits in the animal tree of life are characters with exceptionally low preservation potential, i.e.: cytological and histological innovations. Apomorphies with higher preservation potential do appear early in the fossil record, and support the affiliations of certain controversial fossil taxa. Overall, our results paint a picture of a late Proterozoic world that was probably dominated by small and simple stem members of major metazoan groups: animals that would resemble extant marine invertebrate larvae, placozoans and aspiculate sponges. This study sheds light on the late Proterozoic diversification of animals, and aims to inform future work on exceptional sites such as Doushantuo about what early animal fossils could be like.
The Cadomian arc and orogeny in SW Iberia revisitedTOP
C. Quesada1
1Instituto Geológico Minero de España, Madrid, Spain. Email: quesada.cecilio@gmail.com
The Ossa Morena Zone (OMZ) of SW Iberia records a complex pre-Variscan geodynamic evolution including the following major events: (i) subduction/arc growth (~630-550 Ma); (ii) back-arc opening and spreading (~600-570 Ma); (iii) back-arc closure, polyphase deformation (Cadomian orogeny) and growth of a secondary arc at the N margin of OMZ (~565-535 Ma); and (iv) subduction halt and rapid transition to rifting (~535-530 Ma).
Neoproterozoic subduction/arc development at/near the outer margin of N Gondwana characterizes most Peri-Gondwanan terranes. In the SW Iberia segment no basement exposures occur but an Andean-type context is suggested by Ediacaran mature sandstones and carbonates (Serie Negra succession). Detrital and inherited igneous zircon data link this part of Cadomian Arc to the West African craton. Opening of a back-arc basin is indicated by tholeiitic mafic/ultramafic sequences interpreted as ophiolites, e.g. Calzadilla de los Barros (gabbro protolith dated at ca. 600 Ma). Back-arc basin opening and spreading was probably related to slab-rollback, whereas its inversion may have been connected to either subduction rate increase, a decrease of the subduction angle or both. Inversion was probably accommodated by antithetic subduction of the basin oceanic lithosphere and secondary arc growth (Malcocinado volcanics and associated plutons). Significant sinistral strike-slip components are suggested by juxtaposition of the OMZ to units derived from more easterly N-African areas (Saharan metacraton/Arabian-Nubian shield), currently outcropping north of the OMZ. Formation of a wide foreland basin onto those units, incorporating detritus sourced in the OMZ, and location of the secondary arc rocks close to the northern OMZ boundary support the antithetic polarity of subduction that led to closure of the back-arc basin. Finally, a process of oblique ridge-trench collision is envisaged as responsible for a rapid shift from subduction to rifting (Baja California style), which culminated in opening the Rheic Ocean ca. 495 Ma.
Phosphorites as a new window to early Cambrian evolution of sponges: Fontanarejo, central Spain and basal Niutitang Formation, Hunan, ChinaTOP
J. Reitner1,2, C. Luo3,4
1Department of Geobiology, Centre of Geosciences of the University of Göttingen Goldschmidtstraße 3, Germany. Email: jreitne@gwdg.de
2Academy of Science and Humanities, Theater Str. 7, 37077 Göttingen, Germany.
3CAS Key Laboratory of Economic Stratigraphy and Palaeogeography, Nanjing Institute of Geology and Palaeontology (LESP, NIGPAS), 39 East Beijing Road, China.
4Center for Excellence in Life and Paleoenvironment, Chinese Academy of Sciences, China.
Investigations of the early evolution of sponges are impeded by the dearth of fossil records. Despite of the questionable Ediacaran sponge-like fossils, the earliest convincingh sponge spicules appeared from ca. 535 Ma. However, disarticulated spicules are of little taxonomic value, and almost all the articulated and well preserved sponge fossils were reported after the beginning of Cambrian Age 3, from Burgess-Shale-type preservation. Here we demonstrate three-dimensionally preserved articulated sponge fossils from the lower Cambrian phosphorites in South China (the basal Niutitang Formation) (Figs. 15, 16) and from Fontanarejo in Central Spain (Fig. 14). The ages of both fossil horizons are no younger than the Cambrian Age 2 (Álvaro et al., 2016; Luo & Reitner, 2019).
|
 Figure 14.—Partly articulated hexactinellid sponge embedded in brownish apatite from Fontanarejo (Central Spain). The only spicules are hexactins in different sizes. Figure 14.—Partly articulated hexactinellid sponge embedded in brownish apatite from Fontanarejo (Central Spain). The only spicules are hexactins in different sizes.
|
|
|
 Figure 15.—Main phosphatic sponge layer from the base of the Niutitang formation below the Mo-Ni ore layer in Sancha (China). Figure 15.—Main phosphatic sponge layer from the base of the Niutitang formation below the Mo-Ni ore layer in Sancha (China).
|
|
|
 Figure 16.—Detailed preservation of a phosphatized hexactinellid sponge from the main phosphatic sponge layer in Sancha. Spicules are in place and exhibit various sizes, including micro-hexactins. Figure 16.—Detailed preservation of a phosphatized hexactinellid sponge from the main phosphatic sponge layer in Sancha. Spicules are in place and exhibit various sizes, including micro-hexactins.
|
|
The Niutitang sponges are pre-served as iso-lated, oval bodies in nodular, carbo-naceous cherty phosphorites (Fig. 15). Petrographic observation shows that these phosphorites were dominantly formed by authigenic precipitation. Spicules in these concretions are either completely embedded in cryptocrystalline phosphate, or immediately overgrown by an early isopachous phosphate cement, leaving the remaining interspace and filled by other components (Luo & Reitner 2019). The specimens from Fontanarejo were found in re-deposited phosphatic pebbles, but with their original preservation similar to that of the Niutitang fossils (Reitner et al., 2012). Thin section observation and 3-D reconstruction confirm that these spicules represent sponge skeletal frames preserved in situ.
The taxonomic diversity of the-se fossil com-munities has not yet been fully explored. Two sorts of skeletal frames have been re-cognized in the Niutitang materials: one is composed of hexactines in at least three size-hierarchies; the other is dominated by diactines and few hexactines which show perpendicular arrangement in 3-D space. These fossils are here interpreted as stem group Hexactinellida. Apart from this, some of the disarticulated spicules scattered in the matrix are identical to typical demosponge spicules. Meanwhile, Saetaspongia-like skeletal frame and frame consisting of fused hexactines have been recognized from the Fontanarejo materials.
These observations indicate that the lower Cambrian phosphorites can be a new taphonomic window to study the early evolutionary history of sponges. Further taxonomic and geochemical analyses are required and ongoing to reveal more details.
Dynamic redox changes in shallow seawater environments following the Marinoan glaciationTOP
A.S. Rodler1,2, S.V. Hohl3, C. Gaucher4, G.J.B. Germs5, W. Hegenberger6†, R. Frei2, S. Goderis1, P. Claeys1
1Department of Chemistry, Research Group of Analytical, Environmental and Geo-Chemistry, Vrije Universiteit Brussel, Brussels, Belgium. Email: Alexandra.Rodler@vub.be
2Department of Geosciences and Natural Resource Management, Section of Geology, University of Copenhagen, Copenhagen, Denmark.
3State Key Laboratory of Marine Geology in the School of Ocean and Earth Science, Tongji University, Shanghai, China.
4Institute of Geological Sciences, University of the Republic, Montevideo, Uruguay.
5Geology Department, University of the Free State, Bloemfontein, South Africa.
6Bergstrasse 24, Windhoek, Namibia.
The Neoproterozoic Era witnessed dramatic environmental changes with global low latitude glaciations in the Cryogenian Period, followed by a significant rise in environmental oxygenation. These events may have set the stage for the evolution of first multicellular organisms in the Ediacaran Period. However, the biogeochemical changes that occurred in post-glacial marine environments need to be constrained in more detail before we can evaluate the implications for the evolution of early life.
To elucidate the environmental conditions following the Marinoan glaciation (ca. 635 Ma) we performed a series of geochemical analyses in carbonate lithologies from the Cryogenian to Edicaran Witvlei Group, Namibia (Prave et al., 2011): δ13C and δ18O values , major- and trace elements were obtained on carbonate leachates followed by the analysis of stable Cr isotope compositions. Due to its redox-sensitivity, this isotope system holds the potential to record past fluctuations in oxygen levels in the atmosphere-hydrosphere system. Furthermore, no significant isotope fractionation is expected for natural carbonates, which renders ancient carbonates as promising archives for Cr-isotope-based paleo-redox reconstructions.
This study presents data obtained in post-Marinoan cap dolostones, low Mg dolostones and limestones hosting abundant in situ domical and columnar stromatolites and their clasts (Hegenberger, 1993). The strata were deposited in intertidal and shallow subtidal/shelf lagoon settings with recorded shale normalized (SN) REY patterns similar to those of modern oxidized seawater with relative enrichments of heavy over light rare earth elements (PrSN/YbSN < 1). Further, distinct negative Ce anomalies (CeSN/Ce*SN down to 0.5) and superchondritic Y/Ho ratios (up to 73) have been observed. Carbonate δ13C values are characteristically ‘light’ (-2 to -5 ‰) and show no correlation with δ18O or Sr/Mn ratios, arguing against a significant diagenetic fluid overprint. Trace element concentrations, such as those of Fe, V and Cr show variable but elevated levels throughout the studied sections. First Cr-isotope data of these carbonates fluctuate from negative to positive values relative to bulk silicate Earth (BSE; Schoenberg et al., 2008), particularly for post-Marinoan intertidal and subtidal settings with abundant aragonite fans (-0.2 to +0.3‰), while shallow subtidal settings with abundant lithified stromatolites show pronounced positively fractionated δ53Cr values of up to +0.7 ‰. The observed δ53Cr fluctuations indicate sufficiently high oxygen levels to oxidize and mobilize Cr during weathering processes on land following the Marinoan glaciation, as well as sufficiently oxygenated seawater settings to stabilize these positively fractionated Cr-isotope values. In summary, our results reveal dynamic redox fluctuations in shallow marine settings that were influenced by abundant biological activity following the Marinoan glaciation.
Geochemical and isotopic (Sm-Nd) evidence for two contrasting Late Ediacaran successions in SW Iberian MassifTOP
E. Rojo-Pérez1, R. Arenas1, J.M. Fuenlabrada2, S. Sánchez Martínez1, R. Díez Fernández3, L.M. Martín Parra4, J. Matas4, A.P. Pieren3
1Departamento de Mineralogía y Petrología e Instituto de Geociencias (UCM, CSIC). 28040 Madrid, Spain. Email: e.rojo@ucm.es
2Unidad de Geocronología (UCM). 28040 Madrid, Spain.
3Departamento de Geodinámica, Estratigrafía y Paleontología (UCM). 28040 Madrid, Spain.
4Instituto Geológico y Minero de España. 28760 Tres Cantos, Madrid, Spain.
The Cadomian basement in central and southern Europe is composed by Late Ediacaran-Cambrian rock complexes that appear dismembered as part of different geotectonic zones along the Variscan Orogen, formed in Devonian and Carboniferous times during the main stages of the Pangea assembly. The stratigraphic record of these ancient series contains geochemical and isotopic clues about the origin and evolution of different basins along the Gondwana margin, whose current location does not correspond in some cases with their original position on the African paleo-margin. The southern branch of the Variscan Orogen, the Iberian Massif, contains an excellent stratigraphic record of Ediacaran and Paleozoic sedimentary series in addition to several sutures generated during the long convergence and collision between Gondwana and Laurussia. The nature of the boundary between an autochthonous domain (Central Iberian Zone) and the allochthonous domains (Ossa-Morena Complex) in the SW of Iberian Massif, is under discussion. Previous interpretations, based on U-Pb geochronology of detrital zircons, have suggested that the Ediacaran and Early Paleozoic sedimentary series from the Central Iberian Zone were deposited in more eastern locations along the Gondwana margin than the equivalent series of the Ossa-Morena Complex. However, whole rock geochemistry and Sm-Nd isotope sources of the contemporary siliciclastic series of the Central Iberian Zone and the Ossa-Morena Complex have not yet been compared, bearing in mind they allow a reliable time-resolved comparison between different crustal sectors. The present research compares the whole rock geochemical data and Sm-Nd isotopic tracers of two analogous Ediacaran siliciclastic series (c. 565 Ma), located on both sides of the Central Iberian Zone – Ossa-Morena Complex boundary. The comparison is established for very close domains of both regions: the southernmost sector of Central Iberian Zone, including only the Lower Unit of the so-called Schist and Greywacke Complex (Lower Alcudian), and the Serie Negra Unit (Black Series) of the northernmost part of the Ossa-Morena Complex, designated as the Obejo-Valsequillo Domain.
The Serie Negra Unit has traditionally been divided into two formations, from bottom to top: the Montemolín Formation, whose maximum depositional age has been estimated at c. 590 Ma, and the Tentudía Formation (c. 565-541 Ma). With the exception of the Mérida Massif, the exposed section of the Serie Negra within the Obejo-Valsequillo Domain belongs to the upper part of the Tentudía Formation, and is composed by metasandstones, volcanogenic metagreywackes, slates and phyllites, black quartzites, metacherts and layers of micaschists and limestones. Major and trace geochemical data of the Serie Negra and Lower Alcudian siliciclastic rocks (mainly metagreywackes) are consistent with an immature character of the samples and a limited alteration in the source area. All the study samples follow similar Post Archean Australian Shale and Upper Continental Crustal patterns, suggesting an igneous provenance. Likewise, their geochemical features suggest an active margin setting as the most probable context for the deposition of both Ediacaran sedimentary series. Nevertheless, the Sm-Nd composition of the Serie Negra shows a range of higher negative εNd(0) and εNd(565) values (-9.8 to -20.5 and -4.0 to -13.9, respectively), relative to the Lower Alcudian values (-6.4 to -8.8 and -1.4 to -3.0, respectively). TDM model ages of the Serie Negra range from Mesoproterozoic to Paleoproterozoic (1421-2040 Ma) and are significantly older than those of the Lower Alcudian series (1256-1334 Ma). These isotopic results would situate the source areas of the Serie Negra in the periphery of a cratonic region, likely in the realm of the West African Craton. This old cratonic isotope sources would be mixed with other younger sources, probably derived from the nearby Ediacaran magmatic arc. However, the different Nd model ages of the Lower Alcudian indicate source areas isotopically younger than those for the Serie Negra, which would suggest deposition at distant eastern sections along the Gondwana margin.
According to these data, the contrasting Sm-Nd isotopic sources make it hard to sustain that the siliciclastic rocks of the Lower Alcudian and Serie Negra were deposited close to one another. The current boundary between the Central Iberian Zone and Ossa-Morena Complex (Obejo-Valsequillo Domain) may represent a zone with significant tectonic transport. A major Variscan thrusting developed in a dextral convergence setting can achieve the juxtaposition of distant terranes, as required for the Ediacaran series along this boundary. A similar approach could be likely investigated in other areas with similar geology along the Variscan Orogen, where there are available data to test the existence of major tectonic boundaries in the absence of clear structural and petrological indicators.
Following the logic behind biological interpretations of the Ediacaran biotasTOP
B. Runnegar1
1Department of Earth, Planetary and Space Sciences and Molecular Biology Institute, University of California, Los Angeles, CA 90095-1567, USA, runnegar@ucla.edu
For almost 150 years, megascopic structures in siliciclastic sequences of terminal Precambrian age have been frustratingly difficult to characterize and classify. As with all other areas of human knowledge, progress with exploration, documentation and understanding is growing at a superexponential rate. Nevertheless, there is much to learn from following the evolution of the logic behind the biological interpretations of these enigmatic fossils. In this presentation I review five long-established taxa that represent core components of widely recognized clades that are still difficult to graft onto the tree of life. These “orphan plesions” occupy roles that were once dominated by some famous former Problematica such as archaeocyaths, graptolites and rudist bivalves. In some of those cases, taxonomic enlightenment was brought about the discovery of new characters; in others it required a better knowledge of their living counterparts. Can we use these approaches to rescue the orphans? The five taxa that will be examined in this way are Aspidella (Arboreomorpha), Dickinsonia (Dickinsoniomorpha), Ernietta (Erniettomorpha), Kimberella (Kimberberellomorpha) and perhaps Archaeichnium (Problematica).
Microfossils from the Doushantuo Formation at Liujing in South China, with implications for early-mid Ediacaran biostratigraphyTOP
X. Shang 1,2, P. Liu 1, M. Moczydłowska2
1Institute of Geology, Chinese Academy of Geological Sciences, Beijing 100037, China. Email: shangxiaodong13@mails.ucas.ac.cn
2Department of Earth Sciences, Uppsala University, Uppsala 75236, Sweden.
The distribution of the Ediacaran acritarchs in South China is largely controlled by preservation factors and restricted to the presence of phosphorites and early diagenetic chert nodules, and it shows significant palaeogeographic heterogeneity (Jiang et al., 2011; Muscente et al., 2015). The microfossils in chert nodules and phosphatized microbiotas were mainly recorded in many sections located in the depositional settings ranging from the inner shelf, shallow-water margins of lagoon, over the shelf margin and to the upper slope facies (Chen & Liu, 1986; Zhou et al., 2002; Liu et al., 2014b; Xiao et al., 2014b; Hawkins et al., 2017; Ouyang et al., 2017; Liu & Moczydłowska, 2019). However, fossiliferous chert nodules have not been previously reported from the successions at the center of lagoon. In this study, we present a diverse microfossil assemblage of the Ediacaran acanthomorphic and sphaeromorphic acritarchs, multicellular algae, and filamentous cyanobacteria from the Liujing succession in the Songlin area, which was deposited near the center of intrashelf lagoon (Jiang et al., 2011). The Liujing assemblage extends the palaeographic distribution of Ediacaran acritarchs and provides new useful materials for subdivision and regional correlation of Ediacaran strata in South China.
The microfossils in the Liujing section represent taxonomically identified 25 genera, 44 species, and one undetermined species. The assemblage is dominated by Cymatiosphaeroides forabilatus and Mengeosphaera membranifera sp. nov.. It also shares many species with other assemblages in the Doushantuo Formation of the Yangtze Gorges area, Weng’an, Siduping, Lujiayuanzi, Chaoyang in South China, and coeval microbiotas from Australia, Siberia, the East European Platform, India, Mongolia and Svalbard, showing a high taxonomic similarity to other regional/global microfossil assemblages (Knoll, 1992; Moczydłowska et al., 1993; Tiwari & Knoll, 1994; Zhou et al., 2002; Grey, 2005; Vorobeva et al., 2009; Liu et al., 2014b; Xiao et al., 2014b; Hawkins et al., 2017; Ouyang et al., 2017; Anderson et al., 2019; Liu & Moczydłowska, 2019).
As the representative species of Tianzhushania spinosa biozone recognized in the member II of Doushantuo Formation in the Yangtze Gorges area, T. spinosa does not occur in the Liujing microfossil assemblage, suggesting that the assemblage range is above the range of T. spinosa. It is likely that the stratigraphic position of the Liujing assemblage does not pertain to the lower Doushantuo Formation, or at least above the LAD of T. spinosa. It is also supported by the relatively high stratigraphic intervals containing microfossils at this entire geological succession. The Liujing assemblage contains Tanarium conoideum and Cavaspina basiconica which are the eponymous species of the third Assemblage Zone recognized in the Yangtze Gorges area of South China (Liu and Moczydłowska, 2019). It also has many other species in common and therefore may be assigned to this biozone. The Liujing assemblage does not contain the representative species of the overlying Tanarium pycnacanthum–Ceratosphaeridium glaberosum Assemblage Zone, suggesting that its biostratigraphic position may lie below the lower boundary of the fourth zone and possibly coincides with the barren interval. If this is the case, the Liujing assemblage could also be in part correlated with the stratigraphic interval that is equivalent to the sedimentary gap between the third and overlying Tanarium pycnacanthum-Ceratosphaeridium glaberosum Assemblage Zone.
The Ediacaran-Cambrian transition: the emerging record from Small Carbonaceous Fossils (SCFs)TOP
B.J. Slater1, T.H.P. Harvey2, R. Guilbaud3, S. Willman1, G.E. Budd1, N.J. Butterfield4.
1Department of Earth Sciences (Palaeobiology), Uppsala University, Villavägen 16, SE-75236 Uppsala, Sweden. Email: ben.slater@geo.uu.se
2School of Geography, Geology and the Environment, University of Leicester, Leicester LE1 7RH, UK.
3Lancaster Environment Centre, Lancaster University, Lancaster, LA1 4YQ, UK.
4Department of Earth Sciences, University of Cambridge, Downing Street, Cambridge, CB2 3EQ, UK.
The dramatic changes in the fossil record across the Ediacaran–Cambrian transition seemingly capture the initial diversification of bilaterian animals. This radiation is principally recognised by the signal from mineralized skeletal fossils (Budd & Jensen, 2003; Butterfield, 2007), along with the conspicuous emergence and diversification of complex trace fossils (Jensen, 2003). However, the existing record from shelly fossils captures only a small fraction of the original diversity, as revealed at Konservat-Lagerstätten sites such as the Burgess Shale, where the vast majority of fossilised taxa produce no mineralized skeletal parts (Butterfield, 2003). Despite the exceptional snapshots afforded by such Lagerstätten sites, their inherent rarity limits their contribution to tracing the overall patterns of the Ediacaran-Cambrian transition and subsequent Cambrian radiation (Butterfield, 2003; Slater et al., 2018a). In particular, Burgess Shale-type Lagerstätten sites are noticeably absent from near to the Ediacaran-Cambrian boundary, and from the earliest Cambrian Terreneuvian Series (~541-521 Ma). Consequently, our knowledge of unmineralized biota from this critical timespan is severely impaired. Recently, the newly identified record of ‘small carbonaceous fossils’ (SCFs) has emerged as an alternative means of detecting and tracking unmineralized taxa, otherwise absent from the conventional shelly fossil record (Butterfield & Harvey, 2012). SCFs comprise a polyphyletic assortment of organic remains sourced from metazoans, protists, algae and other organisms. While SCFs typically preserve only incomplete flattened fragments of the original organism, these can frequently capture exquisitely detailed cuticular components and distinctive microstructures that betray the affinity of their producer, even at the specific level (e.g., Harvey et al., 2012; Smith et al., 2015).
A series of new SCF biotas identified from Ediacaran–Cambrian successions on Baltica (Slater et al., 2017, 2018a; Slater & Willman, 2019), Laurentia (Slater et al., 2018b) and other regions is adding a new dimension to the fossil record across this transition (Fig. 17, 18). Initial investigations have shown that the distribution of oxygen was a major factor in determining the distribution of animals and preservation of SCFs (Guilbaud et al., 2018). Taken together, these studies have also revealed a characteristic temporal signal: early Cambrian (Stage 3-4) SCF assemblages produce a rich diversity of recognisable metazoan remains, many of which are assignable to Cambrian taxa known from Burgess Shale-type Lagerstätten elsewhere. Earliest Cambrian (Terreneuvian) SCF assemblages push back the records of several major groups, yet these older assemblages tend to contain only a moderate diversity of metazoans usually dominated by spines with simple morphologies. However, the fine-scale characteristics of these spines including their shape, texture and thickness can distinguish different categories of metazoans even here. The abundance of ‘large’ benthic non-animal SCFs in the Fortunian is further evidence for the persistence of widespread microbial matground-type substrates in the earliest part of the Cambrian. Preliminary investigations of latest Ediacaran SCF assemblages reveal a largely protistan-dominated record, including evidence for multiple boundary-crossing elements of the assemblage. In spite of the substantial differences in mode of preservation and taxonomic coverage, the pattern emerging from this new SCFs record is therefore one that still coarsely resembles that recognised among the more conventional records of mineralized hard-parts and trace fossils.
|
 Figure 17.—Metazoan SCFs from the early Cambrian of Baltica. A-B. Simple spinose elements; C–D. Priapulid-derived cuticular sclerites; E. Palaeoscolecid plate; F. Sclerite sourced from a Wiwaxia. A, B, from the Fortunian of western Finland. C-F, from the early Cambrian of southern Sweden. Scale bars: A-E = 100 µm; F = 100 µm. Figure 17.—Metazoan SCFs from the early Cambrian of Baltica. A-B. Simple spinose elements; C–D. Priapulid-derived cuticular sclerites; E. Palaeoscolecid plate; F. Sclerite sourced from a Wiwaxia. A, B, from the Fortunian of western Finland. C-F, from the early Cambrian of southern Sweden. Scale bars: A-E = 100 µm; F = 100 µm.
|
|
|
 Figure 18.—Assortment of non-animal SCFs from the earliest Cambrian of Baltica. A. Large acritarch (Lontohystrichosphaera grandis), reminiscent of forms known from Proterozoic strata; B. Large sheet-like problematica (Retiranus balticus); C. Filamentous Polythrichoides-like SCF; D. Problematic annulated tubular fossil (Sokoloviina costata); E. Large Leiosphaeridia; F, G. Palaeolyngbya-and Rugosoopsis-like pseudo-segmented filaments. A–D, G, from the Fortunian of Estonia; E, F, from the Fortunian of western Finland. Scale bar: A, C–G = 200 µm; B = 400 µm. Figure 18.—Assortment of non-animal SCFs from the earliest Cambrian of Baltica. A. Large acritarch (Lontohystrichosphaera grandis), reminiscent of forms known from Proterozoic strata; B. Large sheet-like problematica (Retiranus balticus); C. Filamentous Polythrichoides-like SCF; D. Problematic annulated tubular fossil (Sokoloviina costata); E. Large Leiosphaeridia; F, G. Palaeolyngbya-and Rugosoopsis-like pseudo-segmented filaments. A–D, G, from the Fortunian of Estonia; E, F, from the Fortunian of western Finland. Scale bar: A, C–G = 200 µm; B = 400 µm.
|
|
Age constraints for the Ediacaran-Cambrian boundary in the Iberian Chains, NE SpainTOP
M. Streng1
1Uppsala University, Department of Earth Sciences, Palaeobiology, Villavägen 16, 75236 Uppsala, Sweden. Email: michael.streng@geo.uu.se
The Paracuellos Group is the oldest sedimentary sequence of the Iberian Chains in north-east Spain. The group is subdivided into three formations which are, in stratigraphic order, the Sestrica Formation, known only from drill cores, the Saviñan Formation and the Aluenda Formation (Álvaro & Blanc-Valleron, 2002). The sequence has traditionally been considered as Neoproterozoic in age based on its stratigraphic position (e.g., Lotze, 1961), the occurrence of the trace fossil Torrowangea aff. rosei Webby in the middle part of the Saviñan Formation (Liñan & Tejero, 1988), and putative fossils of Cloudina Germs in the Aluenda Formation (Streng, 1996; Álvaro & Blanc-Valleron, 2002).
The Paracuellos Group is exposed in two geographically separated areas where it forms the core of faulted anticlines, the Paracuellos antiform and the Codos antiform in the in the northwestern and central Eastern Iberian Chains, respectively (Álvaro & Blanc-Valleron, 2002). In the Paracuellos antiform, the boundary between the Paracuellos Group and the overlying Bámbola Formation is represented by an erosive contact and represents a hiatus (Liñan & Tejero, 1988; Álvaro & Blanc-Valleron, 2002). In contrast, in the Codos antiform a more gradual transition between the two formations can be observed (Streng, 1996; Álvaro et al., 2002). The Ediacaran-Cambrian boundary was traditionally placed at the boundary between the Aluenda Formation and the Bámbola Formation.
The putative Cloudina fossils have been recovered from a carbonate bed that forms the base of the Aluenda Formation in the Codos antiform, termed the Codos Bed by Álvaro & Blanc-Valleron (2002). However, already at the time of discovery, Streng (1996) pointed out that the tube-shaped fossils lack the characteristic cone-in-cone structure of Cloudina. In addition, cross-sections of a variety of other tube-like small shelly fossils had been observed within the carbonate bed, partly exhibiting internal septate structures and/or external ornamentation (Streng, 1996), and indicating an unusual diversity of shelly fossils for the Neoproterozoic. Similar observations were also made by Álvaro et al. (2016) who placed the Aluenda Formation with the Codos Bed questionably in the early Cambrian (Tommotian). Gámez Vintaned et al. (2018) re-studied the section (their ‘Codos formation’) and also observed a variety of shelly fossils within the carbonate bed. They reported the presence of at least seven genera, including taxa such as Cupitheca Duan, Cobboldiella Kerber, and Pseudorthotheca Cobbold as well as halkieriid sclerites. In addition, a new genus and species with a tube-shaped shell was introduced, Codositubulus grioensis. Despite the rich fossil content and the identification of common early Cambrian taxa, Gámez Vintaned et al. (2018) argue for a late Ediacaran or earliest Cambrian age for the association. One of their reasons for this age is the supposed absence of molluscan shelly fossils in the Codos Bed.
Re-investigation of crack-out material of the Codos Bed, however, has revealed the presence of also molluscan taxa. Poorly preserved composite(?) molds of two helcionellid taxa were recovered, which tentatively have been assigned to Anabarella cf. plana Vostokova and Igorella sp. Specimens are rare though and each taxon is represented by a single specimen only. Nevertheless, they unambiguously indicate a Terreneuvian age (late Fortunian to Cambrian Age 2) of the Codos Bed and confirm the stratigraphic position proposed by Álvaro et al. (2016). In combination with the recent fossil findings by Gámez Vintaned et al. (2018), a previously suggested correlation of the Codos Bed with the Heraultia Limestone of southern France also gains additional support (Álvaro et al., 2010, 2016). The fossils initially described as Cloudina? sp. by Streng (1996) are currently under scrutiny and are considered to represent small shelly fossils characteristic for the lowermost Cambrian, potentially similar to forms such as Pseudorthotheca or Tiksitheca Missarzhevsky, rather than Cloudina. Also, their affinity with the new taxon Codositubulus will be tested.
In conclusion, the newly discovered fossils from the Codos Bed confirm that the Paracuellos Group is not in its entirety Neoproterozoic in age but that it is in part early Cambrian. The Ediacaran-Cambrian boundary is stratigraphically below the Aluenda Formation, i.e. within the 300 to 400 m thick sequence of the two underlying formations. Paleontological re-investigation of the sandstones and shales of especially the Saviñán Formation, as well as re-evaluation of the stratigraphic significance of Torrowangea aff. rosei, are required to further constrain the position of the boundary.
Infaunal augurs of the Cambrian Explosion: An Ediacaran trace fossil assemblage from Mount Dunfee, Nevada, USATOP
L.G. Tarhan1, P.M. Myrow2, E.F. Smith3, L.L. Nelson3, P.M. Sadler4
1Department of Geology and Geophysics, Yale University, New Haven, CT 06520, USA. Email: lidya.tarhan@yale.edu
2Department of Geology, Colorado College, Colorado Springs, CO 80903, USA.
3Department of Earth and Planetary Sciences, Johns Hopkins University, Baltimore, MD 21218, USA.
4Department of Earth and Planetary Sciences, University of California, Riverside, Riverside, CA 92521, USA.
The evolution of infaunal animals dramatically modified Earth’s surface environments. Bioturbating (sediment-mixing) infaunal animals are preeminent ecosystem engineers that, by physically and chemically reworking seafloor sediment, shape nutrient cycling and the structure of benthic communities on both local and ocean-wide scales. Given their substantial environmental and ecological impact in modern marine settings, constraining the timing of the emergence and radiation of infauna is an essential step in reconstructing the coevolution of ancient life and environments.
The earliest sediment-displacement structures (i.e., trace fossils) created by bilaterian animals, simple curvilinear or meandering furrows, first appear in upper Ediacaran strata and record the advent of the infaunal lifestyle (e.g., Jensen et al., 2006). However, the majority of these structures are small, morphologically simple, not significantly sediment-penetrative and, although occasionally locally abundant, relatively uncommon by the standards of Phanerozoic trace fossil assemblages (e.g., Jensen et al., 2006). Uppermost Ediacaran strata record more complex styles of infaunalization, including burrows of full-relief, spiraling, sinusoidal, zig-zag, serial, plug, U- or J-shaped morphology (e.g., Buatois et al., 2018; Chen et al., 2013; Darroch et al., 2016; Gehling et al., 2001; Högström et al., 2013; Jensen et al., 2000; Jensen & Runnegar, 2005; Linnemann et al., 2019; McIlroy & Brasier, 2017; Oji et al., 2018; Parry et al., 2017). These uppermost Ediacaran trace fossils record the pre-Phanerozoic presence of burrowing macrofaunal and meiofaunal bilaterians and the advent of complex infaunal strategies typically associated with Cambrian metazoans. However, these more complex trace fossil morphologies are, in Ediacaran successions, relatively rare - reports of these structures are commonly confined to a single specimen, bedding plane or locality (reviewed in Tarhan et al., 2018); not until the early Phanerozoic do assemblages of morphologically and ethologically complex trace fossils become relatively ubiquitous. Additionally, the majority of these Ediacaran-aged Cambrian-style structures occur only meters to tens of meters below the Ediacaran-Cambrian boundary.
We present a new trace fossil assemblage, recording complex, Cambrian-style infaunalization, from the upper Ediacaran Dunfee Member of the Deep Spring Formation at Mount Dunfee, Nevada, more than 500 m below the local Ediacaran-Cambrian boundary, as previously defined at the first occurrence of Treptichnus pedum, and nearly 400 meters below the nadir of an excursion proposed to correlate to the basal Cambrian negative carbon isotope excursion (BACE) (Corsetti & Hagadorn, 2003; Smith et al., 2016). The Ediacaran succession at Mount Dunfee contains a variety of trace fossils, including simple meandering furrows (Helminthoidichnites, Helminthopsis) and shallowly emplaced infilled burrows (Planolites), as well as regularly sinuous structures (Cochlichnus), which are preserved along bedding planes of the heterolithic lower Dunfee Member of the Deep Spring Formation (Gevirtzman and Mount, 1986). Additionally, the Dunfee Member trace fossil assemblage includes structures of more complex morphology, including treptichnids, as well as zigzagging and shallowly sinuous or spiraling forms similar to trace fossils described as Belorhaphe and Helicolithus, respectively, from uppermost Ediacaran strata of Finnmark (Högstrom et al., 2013; McIlroy & Brasier, 2017). The Dunfee Member also contains bed-penetrative dimpled plugs (Bergaueria). These structures are of millimetric diameters and depths and, with the exception of Bergaueria, are not fabric-disruptive; the beds along which they occur are commonly well-laminated and characterized by an ichnofabric index of 1.
The Dunfee trace fossils are unique, among Ediacaran assemblages, in their combination of morphological complexity and stratigraphic distance from the Ediacaran-Cambrian boundary. Statistical sedimentological and chemostratigraphic data indicate that the Dunfee assemblage is one of the oldest documented instances of sediment-penetrative infaunalization, likely predating the Ediacaran-Cambrian boundary by at least a million years. Moreover, these Dunfee Member trace fossils occur along the same horizons as classic upper Ediacaran tubular fossils such as Gaojiashania and Wutubus, suggesting that Ediacaran epifauna and infauna may, in contrast to recent suggestions (e.g., Darroch et al., 2015, 2016), have coexisted and formed ecologically sophisticated communities in proximal shallow marine settings.
Micro-CT scanning reveals possible bed-penetrative burrows from the terminal Ediacaran of southern NamibiaTOP
K. Turk.1, A.T. Cribb2, S.A.F. Darroch1
1Vanderbilt University, Nashville, TN, 37235, USA. Email: katherine.turk@vanderbilt.edu
2University of Southern California, Los Angeles, CA, 90007, USA.
The Ediacaran-Cambrian transition marks one of the most critical geobiological revolutions in Earth history. It is during this period we see not only the first complex macroscopic metazoans in the fossil record, but Earth’s potential first mass extinction as well. The “biotic replacement” hypothesis posits the Ediacara biota were driven to extinction by the evolution of the Cambrian fauna, possibly due to the advent of Cambrian-type ecosystem engineering behaviors such as burrowing. Preliminary micro-CT scans of material from the terminal Ediacaran Spitskopf Formation (Nama Group, southern Namibia) suggest the presence of comparatively large bilaterian burrows, some of which indicate vertical movement. Using CT data and subsequent segmentation, as well as polished thin sections, we identify several unique forms, interpret them in the context of diversity and complexity of bilaterian behaviors, and discuss their potential relevance to broader Ediacaran-Cambrian bioevents.
Late Ediacaran rocks and fossils in the Bodoquena-Pantanal Geopark (West Brazil): Importance and threatsTOP
D.H.G Walde1, A. Silva1, B.L.P. Silva1, A. Soriano2, M.E.A.F. Ramos3
1Federal University of Mato Grosso do Sul, Campus Pantanal, Corumbá, MS CEP 79.304-030, Brazil. Email: walde.detlef@gmail.com
2Bodoquena-Pantanal Geopark, MS, Brazil.
3University of Brasília, Institute of Geosciences, Campus Darcy Ribeiro, Brasília, DF CEP 70.910-900, Brazil.
The Bodoquena-Pantanal Geopark exists since 2009, having a total extension of approximately 40 thousand square kilometers. In this area there are several geosites of different interests, highlighting, in particular, records of support scientific studies regarding the interpretation of the tectono-environmental evolution of the Paraguay Belt (Walde et al., 2015) in the context of global climatic changes during the late Neoproterozoic, as well as discussions about global glaciations and the hypothesis of a Snowball Earth. Still, it focus the understanding of how these glaciations influenced the transition of life evolution among the earliest microbial forms (stromatolites) to more evolved forms, represented by fossils in the region, like Cloudina and Corumbella (Fig. 19 A-C). In the region of the Geopark there are important geological discoveries, such as the Corumbella werneri, Cloudina lucianoi, Cloudina waldei and recently Cloudina carinata, fossils of the Ediacaran Biota, discovered in the limestones along the Paraguay River in the Corumbá region. Corumbella werneri is the oldest mineral-skeletonized fossil ever found in the Americas, dating to about 540 Ma. (Adorno et al, 2017).
|
 Figure 19.—Cloudina lucianoi (Tamengo Formation, Corumbá Group). B. Corumbella werneri (Tamengo Formation, Corumbá Group). C. Tilite with drops-tones (Santa Cruz Formation, Jacadigo Group). Figure 19.—Cloudina lucianoi (Tamengo Formation, Corumbá Group). B. Corumbella werneri (Tamengo Formation, Corumbá Group). C. Tilite with drops-tones (Santa Cruz Formation, Jacadigo Group).
|
|
The region includes an important but fragile ecosystem of broad interest: the swampy Pantanal of Mato Grosso do Sul (Fig. 20A) with a rich wildlife and karst terrains, environments of extraordinary beauty and extreme fragility threatened by any form of occupancy that is not of nature conservation. Other palae-ontological geosites are the dinosaur footprints found in Nioaque and archeolo-gical sites with indigenous pain-tings and ceramics (e.g. Terenas, Kadwéus). There are also historical sites related to the Paraguay War, like the battlefield of Nhandepá, which is an archeological site.
|
 Figure 20.—Typical scenery of the Pantanal region (Mato Grosso do Sul). B. Fe-ore mining areas near Corumbá (Santa Cruz Formation, Jacadigo Group). Figure 20.—Typical scenery of the Pantanal region (Mato Grosso do Sul). B. Fe-ore mining areas near Corumbá (Santa Cruz Formation, Jacadigo Group).
|
|
Potential threats to biodiversity, landscape and cultural heritage in the region of the Geopark (Rolim & Theodorovicz, 2012) are due largely to the misguided tourist exploitation and to the improper management of improvements of agricultural and mining activities (Figure 20B). Other input greatly demanded by the global market is the iron and manganese ores, whose production area of excellence is located around Corumbá, in the massive deposits of Urucum/Jacadigo. In non-metallic mining activities in the Bodoquena Mountain Range the practice of explosives affect the balance of the karst geology, resulting in collapse of caves, burial sinks, irretrievable loss of paleontological material and referential landscape. The advance of agro-pastoral frontier in the Pantanal can configure a threat to the archaeological sites, as well as the biodiversity. The effects of the huge river convoys that navigate the river Paraguay can also represent a risk to archaeological sites, e.g. causing mudslides and water contamination. All these threats could only be controlled through strict compliance with and control of existing legislation. In this sense, the establishment of a Geopark, besides being an initiative to preserve the rich geological and paleontological heritage, has also become an important tool for adequate forms of environmental control, such as land occupation and use.
Cambrian Stage 3 ‘small shelly fossils’ from the Terrades inlier, southeastern Pyrenees, Spain: biostratigraphic and palaeobiogeographic implicationsTOP
E. Wallet1, L. Devaere2, M. Padel3, J.J. Álvaro1, S. Clausen2
1Department of Earth Sciences, Palaeobiology, Uppsala University, Villavägen 16, SE-752 36 Uppsala, Sweden.
2Université de Lille I, CNRS, UMR 8198 - Evo-Eco-Paleo, F-59000 Lille, France. Email: sebastien.clausen@univ-lille.fr
3BRGM, 3 Avenue Claude Guillemin, 45100 Orléans, France.
4Instituto de Geociencias (CSIC-UCM), Dr. Severo Ochoa 7, 28040 Madrid, Spain.
The Pyrenean massif (French-Spanish geographical border) mostly resulted from the convergence between the Iberian and European plates during the Alpine orogenesis. It is structurally subdivided into three zones (the North, Axial and South Pyrenean zones) separated by major W-E-trending thrusts. Most of the Ediacaran-Cambrian successions of the Pyrenees crop out in the Axial Zone, which belongs to the Pyrenean Domain of the Variscan Ibero-Armorican Arc. The Axial Zone has undergone a complex geodynamic evolution which led to fossil depleted, Cambrian metasedimentary successions. As a consequence, the relationships of the Cambrian Pyrenean Domain with neighbouring areas of the northwestern Gondwana margin remain mostly based on: (i) lithostratigraphic correlations, (ii) detrital source analysis through U-Pb laser ablation ICP-MS zircon dating of sedimentary and volcano-sedimentary intervals, and (iii) signature and distribution of magmatic episodes. The Salud/Salut Unit (Gerona Province, Spain), a small inlier of pre-Variscan rocks from the South Pyrenean Zone, makes an exception to this general scheme. In this unit, a ‘lower Cambrian’ succession in the vicinity of the Terrades village has provided the only significant and indisputable Cambrian fossil record of the Pyrenean Chain. This succession mostly consists of shales alternating with reefal and perireefal limestones from which archaeocyaths have been intensively studied, whereas associated skeletonized microfossils remain vaguely mentioned. This study was therefore aimed at describing, for the first time, the small shelly fossil (SSF) assemblage of the Terrades outcrops in order to document the palaeobiogeographic affinities of the Cambrian Pyrenean Domain with surrounding zones of the peri-Gondwanan margin. The studied limestones yielded bradoriids, brachiopods, molluscs, tommotiids (Kelanella altaica) chancelloriids, hyolithelminths, hyoliths and the problematic Rhombocorniculum cancellatum. The recovered assemblage confirms the Cambrian Epoch 2, Age 3 based on associated archeocyath assemblages, although the presence of R. cancellatum allows further international correlations. Moreover, while reported archaeocyaths do not allow to precisely assess the palaeobiogeographic affinities of the Pyrenean Domain, the reported SSF assemblage emphasizes closest affinities with the southern Montagne Noire (Occitan Domain, southern Massif Central, France), as previously suggested by lithostratigraphic and sediment-source analyses. These results would further support an allochthonous origin of the Salud Unit.
The Precambrian-Cambrian boundary at the southeastern margin of the North China CratonTOP
B. Wan1, Q. Tang2, K. Pang1, X. Wang1,3, Z. Bao4, F. Meng1, C. Zhou5, X. Yuan1,3,6, H. Hua4, S. Xiao2
1State Key Laboratory of Palaeobiology and Stratigraphy, Nanjing Institute of Geology and Palaeontology and Center for Excellence in Life and Paleoenvironment, Chinese Academy of Sciences, Nanjing 210008, China. Email: binwan@nigpas.ac.cn
2Department of Geosciences, Virginia Tech, Blacksburg, VA 24061, USA. Email: xiao@vt.edu
3University of Chinese Academy of Sciences, Beijing 100049, China.
4State Key Laboratory of Continental Dynamics and Department of Geology, Northwest University, Xi’an 710069, China.
5CAS Key Laboratory of Economic Stratigraphy and Palaeogeography, Nanjing Institute of Geology and Palaeontology and Center for Excellence in Life and Paleoenvironment, Chinese Academy of Sciences, Nanjing 210008, China.
6Center for Research and Education on Biological Evolution and Environment, Nanjing University, Nanjing 210023, China.
Representatively in Laurentia, the Precambrian-Cambrian boundary has been extensively recognized between the Precambrian basement rock and Cambrian sedimentary cover, which was known as the Great Unconformity and causally linked to the environmental trigger for the Cambrian explosion. A similar unconformity has long been known in the North China Craton (NCC), but its exact stratigraphic location has not been constrained in many successions. Especially reflected in the Huaibei region of the southeastern margin of the NCC, this unconformity has been variously placed beneath or above the Gouhou Formation, which has been interpreted as Tonian, Cryogenian to Ediacaran, or Cambrian in age. To resolve this issue, we carried out an integrated sedimentary petrological, biostratigraphic and detrital zircon geochronological investigation on the Precambrian-Cambrian boundary transitions including the Gouhou Formation and the overlying Houjiashan Formation in the Huaibei region. We confirmed a conglomerate bed within the Gouhou Formation, and the potential Tonian index fossils are preserved in the Lower Member below the conglomerate bed, while all the possible trace fossils, bioturbation-caused mottled structures and the early Cambrian detrital zircons are yielded from the Middle-Upper members above the conglomerate bed. Our data suggest that the Lower Member of the Gouhou Formation is Tonian in age whereas the Middle-Upper members are early Cambrian in age, and the Precambrian-Cambrian boundary lies within the Gouhou Formation and represents a ca. 200-300 Myr depositional gap equivalent to the Great Unconformity. As a result, we propose that the Gouhou Formation be redefined to contain only the lower member, and the middle-ppper members be included in the Houjiashan Formation. Our analysis also confirms that, according to the similar basis in the Huaibei region, the Great Unconformity can be also widely recognized in the Huainan region, southwestern Shandong, eastern Liaoning and southern Jilin provinces along the southeastern margin of the NCC. Additionally, the age distributions of detrital zircons from the Precambrian–Cambrian boundary transitions in Huaibei region provide critical insights into changes of sediment provenance related to the break-up of Rodinia and the final assembly of Gondwana. The recognition of the Great Unconformity in the Huaibei region helps to clarify the tectonic history, basin development, stratigraphic correlation, and paleogeographic reconstruction of the southeastern margin of the NCC during the Tonian-Cambrian periods.
The trilobite biostratigraphy of the Balang Formation, the Duyunian Stages, Qiandong Series (Cambrian Series 2, Stage 4) from Huanglian area in Songtao County, Guzhou Province and its correlation significanceTOP
M.Y. Wu1, Y.L. Zhao2, X.L. Yang2, Z.P. Chen2
1School of economics and management, Guiyang university, Guiyang 550005, China.
2College of Resources and Environment Engineering, Guizhou University, Guiyang 550025, China. Email: zhaoyuanlong@126.com
The Huanglian section, located in the west of Huanglian village, Songtao County, Guizhou Province was the basic section for establishing the Nangaoan Stage and the Duyunian Stage (Peng et al., 2000). We restudy some materials collected from the Balang Formtion of the Huanglian section eighteen years ago. The Balang Formtion in the Huanglian section consists of dark grey and greyish black silty mudstone. Its thickness is 417.8 m. The lithology of the Balang Formation at Huanglian is similar to the Balang Formation in east Guizhou and west Hunan (Zhao et al., 2001).
The trilobites from the Balang Formation of the Huanglian section are very abundant except the lower part, from the base boundary to 38 m. Two trilobite zones are defined according to the collected data: (i) the Oryctocarella duyunensis Zone: this zone occupies the lower part of the Balang Formation from 38 m to 83 m above the base and contains mainly Oryctocarella duyunensis, Duyunaspis duyunensis and Changaspis elongate; Oryctocarella duyunensis makes its first appearance about 38 m above the base of the Balang Formation; and (ii) the Arthricocephlus chauveaui Zone: this zone occupies the middle and upper part of the Balang Formation from 83 m to the upper unit, the “Tsinghsutung Formation”, including Arthricocephlus chauveaui (Peng et al., 2017), A. xinzhaiheensis, Oryctocarella duyunensis (Peng et al., 2017), O. sibirica, O. balangensis, Duyunaspis duyunensis, Changaspis elongata and Redlichia (Pteroredlichia) murakamii. Arthricocephlus chauveaui makes its first appearance at 83 m above the base of the Balang Formation (Figs. 21-22).
|
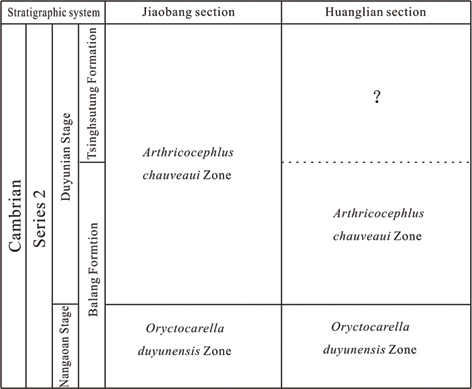 Figure 21.—Correlation chart of the trilobite zonation from Duyunian Stages, Qiandong Series (Cambrian Series 2, Stage 4) in Songtao County and Jianhe County. Figure 21.—Correlation chart of the trilobite zonation from Duyunian Stages, Qiandong Series (Cambrian Series 2, Stage 4) in Songtao County and Jianhe County.
|
|
|
 Figure 22.—A. Arthricocephlus chauveaui Bergeron, 1899, GHL-724-1. B. Oryctocarella balangensis Lu & Qian (in Yin & Li, 1978), GHL-630-1. C. Arthricocephlus xinzhaiheensis Qian & Lin (in Lu et al., 1974), GHL-601-1. D. Oryctocarella duyunensis Qian, 1961, GHL-621-1. E. Redlichia (Pteroredlichia) murakamii Resser & Endo, 1935, GHL-789-1. F. Changaspis elongate Chien, 1961, GHL-725-3. G. Duyunaspis duyunensis Zhang & Qian (in Zhou et al., 1977), GHL-704-1. H. Eosoptychoparia (Danzhaina) conica Yuan (in Zhang et al., 1980a), GHL-857-4; scale bar = 5 mm. Figure 22.—A. Arthricocephlus chauveaui Bergeron, 1899, GHL-724-1. B. Oryctocarella balangensis Lu & Qian (in Yin & Li, 1978), GHL-630-1. C. Arthricocephlus xinzhaiheensis Qian & Lin (in Lu et al., 1974), GHL-601-1. D. Oryctocarella duyunensis Qian, 1961, GHL-621-1. E. Redlichia (Pteroredlichia) murakamii Resser & Endo, 1935, GHL-789-1. F. Changaspis elongate Chien, 1961, GHL-725-3. G. Duyunaspis duyunensis Zhang & Qian (in Zhou et al., 1977), GHL-704-1. H. Eosoptychoparia (Danzhaina) conica Yuan (in Zhang et al., 1980a), GHL-857-4; scale bar = 5 mm.
|
|
The trilobite biostratigraphy of the Balang Formation at the Huanglian section is exactly the same than that of the Balang Formation at the Jiaobang section, which is the potential Global standard stratotype-section and point (GSSP) for the base of the Cambrian Stage 4 (Zhao et al., 2017; Chen et al., 2018) and can correlate with the Henson Gletscher Formation in Freachen land and Peary land of North Greenland (Geyer et al., 2011). The Huanglian section is a good supplementanry section of the Jiaobang section for the Palang Formation.
This study is supported by the Ministry of Science and Technology of China (2015FY310100), and the National Natural Science Foundation of China (No. 41662001 to Wu and Zhao; No. 41772021 to Yang and Zhao).
Was dynamic and heterogeneous redox condition in Ediacaran oceans an environmental impetus for the evolution of bilaterian animal mobility?TOP
S. Xiao1, Z. Chen2, C. Zhou2, X. Yuan2
1Department of Geosciences, Virginia Tech, Blacksburg, Virginia 24060, USA. Email: xiao@vt.edu
2Center for Excellence in Life and Paleoenvironment, State Key Laboratory of Palaeobiology and Stratigraphy, and CAS Key Laboratory of Economic Stratigraphy and Palaeogeography, Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences, Nanjing 210008, China.
Rising oxygen levels are often regarded as an environmental factor facilitating greater biodiversity. Here we consider the possibility that overall low but permissible oxygen levels, and hence dynamic and heterogeneous oceanic redox conditions, may have been a driving force for the evolution of bilaterian animal mobility in the Ediacaran Period.
Phylogenetic relationships among animals indicate that the earliest animals were not actively mobile and the origin of actively mobile bilaterians post-dated the origin of animals . The fossil record shows that the earliest mobile bilaterians appeared in the late Ediacaran Period (ca. 560-540 Ma) , with an apparent increase in ichnofossil diversity and animal mobility in the terminal Ediacaran Period (ca. 550-540 Ma) when many Ediacara-type macro-organisms went extinct . Available geochemical data indicate that the late Ediacaran Period is characterized by a major expansion of marine anoxia relative to preceding and following periods, thus resulting in temporally dynamic and spatially heterogeneous redox conditions in shallow shelf environments where animals lived. Are these environmental and evolutionary events isolated coincidences or causally related? Previous studies were focused on the possibility that environmental stress such as an expansion of oceanic anoxia may have caused extinctions . But is it possible that, while anoxia may have driven some organisms to extinction, it may have also stimulated evolutionary innovations among others?
The observation that Ediacaran trace fossils tend to be closely associated with (presumably cyanobacterial) microbial mats offers insights into the possible relationship between animal mobility and redox conditions. Such an association could be taken as evidence for an early animal behavior to explore localized oxygen oases within cyanobacterial mats in an ocean characterized by relatively low levels of dissolved O2. Because of the dynamic redox conditions in the water column and in the cyanobacterial mat (due to diurnal photosynthesis cycles), animals exploring these oxygen oases must also be able to respond to such dynamic changes by maneuvering in and out the cyanobacterial mats. We hypothesize that such localized and dynamic O2 oases were an environmental stimulus that drove the evolution of mobility among early benthic bilaterian animals.
A new cloudinid from the late Ediacaran of Mongolia and its evolutionary significanceTOP
B. Yang1, M. Steiner2, J.D. Schiffbauer3,4, T. Selly3,4, X. Wu5, C. Zhang1,6, P. Liu1
1MNR Key Laboratory of Stratigraphy and Palaeontology, Institute of Geology, Chinese Academy of Geological Sciences, Beijing 100037, China. Email: benyang@cags.ac.cn
2Department of Earth Sciences, Freie Universität Berlin, Berlin 12249, Germany.
3Department of Geological Sciences, University of Missouri, Columbia, Missouri 65211, USA.
4X-ray Microanalysis Core Facility, University of Missouri, Columbia, Missouri 65211, USA.
5Laboratory of Marine Organism Taxonomy and Phylogeny, Institute of Oceanology, Chinese Academy of Sciences, Qingdao 266071, China.
6School of Earth Science and Engineering, Shandong University of Science and Technology, Qingdao 266590, China.
Ediacaran-Cambrian tubular fossils have been reported from numerous paleocontinents. The cloudinids, particularly, are of significant evolutionary importance because they contain the oldest recognized biomineralizing metazoans. Here we report a new representative of the cloudinids from the Ediacaran strata of western Mongolia. These phosphatic fossils have collars similar to those in Cloudina, but also possess a lamellate wall structure similar to some organic Sabellidites and phosphatic Sinotubulites. Observation of delamination, plastic deformation, and relic content of carbonaceous material suggests that the wall of the new fossil was originally organic. The general morphology and ultrastructure together indicate that the new fossil and potentially other similar cloudinids are comparable to modern siboglinids (Annelida). The shared morphological features in diverse taphomodes among the cloudinids (e.g., Cloudina, Rajatubulus, Saarina, Conotubus, and Multiconotubus) necessitate revision of the cloudinid morphoclade. The interpretation of cloudinids as worm tubes with annelid affinities in the late Ediacaran accords with previous molecular clock estimates of the annelid and molluscan split between 590-540 Ma.
Microfossil assemblage from the Cambrian “Tsinghsutung Formation” of Guizhou, ChinaTOP
X. Yang1, Y. Wang1, Y. Zhao1, B. Wei1, R. Feng1
1College of Resource and Environment Engineering, Guizhou University, Guiyang 550025, China. Email: yangxinglian2002@163.com
The “Tsinghsutung Formation” is an important lithostratigraphic unit, fossiliferous and bearing a potential significance to define the base of Cambrian stage 4. It is well exposed in Balang village, Jianhe County, Guizhou Province, and represents a deposit formed in transition slope belt between the Yangtze platform and the Jiangnan basin, and the overlying Balang Formation, and is overlain by the Kaili Formation, and consisting of thin- to medium-bedded limestone with silty mudstone, as well as fine-grained dolomite in the uppermost part. The microfossils after acetic acid treatment were mainly collected from the dark grey medium-bedded argillaceous limestone in the middle to upper part of “Tsinghsutung Formation”. Microfossil brachiopods in the “Tsinghsutung Formation” are abundant not only quantitatively but also in diversity: 4 genera and 4 species constitute 50 % of the whole microfossil content. The tubular skeletal microfossils also are very common, belonging to hyoliths and arthropods, respectively. Three types of sponge spicules have been found, among which hexactines were dominant. But beyond that, some fragments of trilobites, sponge spicules, echinoderms and algae have also been found during the thin section. This microfossils assemblage is different from the macrofossil assemblage of the “Tsinghsutung Formation” of Jianhe dominated by arthropods. Compared with the microfossil assemblage of coeval strata, the fossil associations from the “Tsinghsutung Formation” share many similarities with the Harkless Formation in southern Nevada because of its low biodiversity, while the diversity of the brachiopod is relatively higher, and most of them have Hadrotreta. The discovery of the microfossils of the “Tsinghsutung Formation” can not only enrich the perfect bio-assemblage of “Tsinghsutung Formation”, but also can provide some fossil evidence to correlate with coeval global strata.
Three-dimensional preservation of early Ediacaran sponge gemmules with cellular resolutionTOP
Z. Yin1, W. Sun1,2, M. Zhu1,3
1State Key Laboratory of Palaeobiology and Stratigraphy, Nanjing Institute of Geology and Palaeontology & Center for Excellence in Life and Paleoenvironment, Chinese Academy of Sciences, Nanjing 210008, China. Email: zjyin@nigpas.ac.cn
2University of Science and Technology of China, Hefei 230026, China.
3College of Earth Sciences, University of Chinese Academy of Sciences, Beijing 100049, China.
As the simplest animals on the planet, sponges have been widely accepted as a monophyletic clade and among the earliest branching lineages of the animal tree. Therefore, the origin and early evolution of sponges are critical for understanding the rise of animals. Molecular clocks estimate that the initial rise of sponges may occur in the Tonian or Cryogenian periods (850-650 Ma), and majority sponge bodyplans or clades probably diversified during Ediacaran. However, the earliest unambiguous sponges did not appear in fossil records until the early Cambrian, about 535 Ma. Even though putative sponge biomarkers have been recovered from Cryogenian and Ediacaran deposits, these molecular fossils have been questioned. Possible sponge grade body fossils have also been reported from Cryogenian and Ediacaran periods, but none of them could be assigned into crown sponges with confidence. Here we present microscopic sponge gemmule-like fossils from the early Ediacaran Weng’an Biota (609 Ma), Guizhou Province, southwest China. These three-dimensionally phosphatized fossils developed a thick, ornamented envelope with a circular micropyle-like opening, and a membrane-bounded multicellular inner body. They show striking similarities to extant sponge gemmules not only in size and morphology, but also in anatomy and histology. This discovery pushes back the oldest fossil record of sponges by at least 77 million years, which is consistent with the molecular clock estimates. Furthermore, in extending the fossil record of crown sponges, this finding also suggests that the earliest stem metazoans and the last common ancestor of metazoans probably arise before the early Ediacaran.
Organic-walled microfossils from sediments of the Cambrian Stage 4 in the Jiaobang Section, eastern Guizhou of ChinaTOP
L. Yin1, K. Wang2, Z. Shen3, Y. Zhao4
1Nanjing Institute of Geology and Palaeontology, Academia Sinica of Sciences, Nanjing 210008, China. Email: leimingyin@aliyun.com
2College of Earth Science and Engineering, Shandong University of Science and Technology, Qingdao 266590, China.
3State Key Laboratory of Biogeology and Environmental Geology, School of Earth Sciences, China University of Geosciences, Wuhan 430074, China.
4College of Resource and Environment Engineering of Guizhou University, Guiyang, China, Guiyang 550025, China.
Previously, early-mid Cambrian organic-walled microfossils were mainly studied and reported from the East European Platform, the Baltoscandian Platform, the southwestern Europe and North America. Many diagnostic morphological forms of organic-walled microfossils had been described. Their biostratigraphic distribution is not only restricted in local region and even known from elsewhere of the world.
The lower Cambrian stratigraphic sequences in the Jiaobang section located at a mountain ridge of eastern Jiaobang village, Jianhe County, eastern Guizhou, South China, include, in ascending order, the Bianmachong, Balang and Tsinghsutung formations with a total of 645m in thickness in the section. 31 morphological genera (including one new genus - Plagasphaera) have been identified from the Balang Formation and the underlying Bianmachong Formation. On the basis of first appearance of diagnostic taxa in the stratigraphic column (Fig. 23), it is noted that several horizons in the stratigraphic column show distinct assemblages. Obviously, the acritarch assemblage from the bed of 82m in distance from bottom of the Balang Formation is more represented the microphytoplankton aspect before the possible geological or ecological change event. This acritarch assemblage can be named as “Celtiberium dedalium-Fimbriagromerella minuta”. It is nearly coincided with alteration of trilobite zones from lower Oryctacarella duyunensis Zone to upper Arthricocephalus chauveaui Zone.
|
 Figure 23.—Geographical locality of the study section, and Jiaobang section with lithostratigraphy and relative trilobite biozones. Figure 23.—Geographical locality of the study section, and Jiaobang section with lithostratigraphy and relative trilobite biozones.
|
|
In addition, at the boundary between the Bianmachong Formation and the overlying Balang Formation, the obvious change of organic-walled microfossil assemblages is also noted, but the gap more far about 24 m in distance from the bottom of the Balang Formation.
A potential trace maker from the Fortunian of South ChinaTOP
X. Zhang1
1State Key Laboratory of the Continental Dynamics, Shaanxi Key Laboratory of Early Life and Environment, Department of Geology, Northwest University, Xi’an, 710069, PR China. Email: xzhang69@nwu.edu.cn
The trace fossil record implies that large worm-like animals were in place along with the skeletonizing organisms during the initial stage of the Cambrian explosion. Body fossils of large worms, however, have so far not been found. The large, soft-bodied, worm-like organism, Vittatusivermis annularius Zhang et al. 2017, from the lowermost Cambrian of South China, which is constrained to the Fortunian Age (541-529 Ma) of the Cambrian Period, provides a potential example of a trace fossil producer. The elongate body of Vittatusivermis was large enough to have supported organ systems and a fluid skeleton that facilitated peristaltic locomotion, thus allowing for more complex patterns of movement than those of flatworms. Its occurrence on the same bedding surface as trace fossils suggests that Vittatusivermis might have produced epichnial trails and shallow burrows on and within sediments. Therefore, Vittatusivermis is likely to have been one of the long expected producers of trace fossils in the earliest Cambrian.
Integrated chemostratigraphic and palaeoclimatic approaches to the subdivision of the Ediacaran SystemTOP
C. Zhou1, S. Xiao2, Q. Ouyang1, C. Guan1, W. Wang1
1CAS Key Laboratory of Economic Stratigraphy and Palaeongeography, Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences, Nanjing 210008, China. Email: cmzhou@nigpas.ac.cn, qouyang@nigpas.ac.cn, cgguan@nigpas.ac.cn, wwang@nigpas.ac.cn
2Department of Geosciences, Virginia Tech, Blacksburg, VA 27061, USA. Email: xiao@vt.edu
The middle Ediacaran Shuram excursion is the most pronounced negative carbon isotopic shift in Earth history. Potential equivalents of the Shuram excursion have been reported in many places around the world (Grotzinger et al., 2011), indicating that it can be used as a potential chemostratigraphic tool for Ediacaran subdivision and global correlation. However, the correlation of those “Shuram equivalents” is based mainly on (i) their pronounced shifting magnitude (e.g., from ~+5‰ to ~-12‰), and (ii) their occurrences in the middle Ediacaran successions loosely constrained by limited palaeontological data. The age of the Shuram excursion is not tightly constrained. The ~ 551 Ma age from South China (Condon et al., 2005) is sometimes cited as directly dating the end of the Shuram excursion, although it should be considered as a minimum age constraint. The onset of the Shuram excursion has not been tightly constrained by radiometric dates either, and estimates range from 580 to 560 Ma.
The Shuram excursion generally shows a simple stratigraphic pattern with a sharp shift to negative δ13C values followed by a slow recovery to positive values (Grotzinger et al., 2011). The δ13C profiles for the upper Doushantuo Formation around the Huangling Anticline, western Hubei Province, South China, however, gives a more complex picture. Whereas only a single δ13C negative excursion occurs in the upper Doushantuo Formation at some sections in the eastern region of the Huangling Anticline (for example, EN3 at the Jiulongwan section), other sections in the western region contain two negative excursions (the lower JNE or the Jiuqunao δ13C negative excursion, and the upper MNE or the Miaohe δ13C negative excursion) separated by a moderately positive excursion (the DPE or the Diaoyapo δ13C positive excursion) (Zhou et al., 2017). Detailed stratigraphic correlation of the DPE and MNE around the Huangling Anticline is still a matter of debate (An et al., 2015); however, it has been widely accepted that only the JNE is correlated with EN3 and the Shuram excursion. Strata hosting the JNE are overlain by a black shale (up to 3 m thick), a dolostone (up to 12 m thick), and the Miaohe Member black shale (up to 23 m thick). The ~551 Ma age was reported for an ash bed in the uppermost Miaohe Member. Considering the typically slow deposition rate of black shales, it is possible that both the onset and the end of JNE (=EN3 or Shuram) were much earlier than 551 Ma.
The ~580 Ma Gaskiers glaciation has been used as a palaeoclimatic marker to subdivide the Ediacaran System at a series level (Narbonne et al., 2012; Xiao et al., 2016). The occurrences of equivalent diamictite of the Gaskiers glaciation are rare in a global scale, and recent geochronological study suggests that a late Ediacaran glacial event occurred at ~565 Ma in West Africa (Linnemann et al., 2018), making the global correlation of the Ediacaran glacial deposition much more challenging than before. However, the ~580 Ma Gaskiers glacial event has great potential as a palaeoclimatic marker in the Ediacaran subdivision, because it temporally subdivides the Ediacaran System into two series of comparable duration (Xiao et al., 2016).
A recent subsidence model analysis of the Ediacaran Johnnie Formation indicated that the Shuram excursion occurred 585-579 Ma and its termination was coincident with the ~580 Ma Gaskiers glaciation (Witkosky & Wernicke, 2018). If the occurrences of Gaskiers glaciation and the Shuram excursion can be proved as globally synchronous events in the future, the combination of chemostratigraphic shift and palaeoclimatic event may provide an ideal criterion to subdivide the Ediacaran System.
ReferencesTOP
| ○ |
Adôrno, R.R.; Do Carmo, D.A.; Walde, D.H.; Denezine, M.; Boggiani; P.C.; Sousa, S.C.; Vasconcelos, J.R.; Tobias, T.C.; Guimarães, E.M.; Vieira, L.C.; Figuereido, M.F.; Caminha, S.A.; Suarez, P.A.Z.; Rodriguez, C.G.V.; Pinho, D.M.; Schneider, G. & Muyamba, R. (2017). Cloudina lucianoi (Beurlen & Sommer, 1957), Tamengo Formation, Ediacaran, Brazil: taxonomy, analysis of stratigraphic distribution and biostratigraphy. Precambrian Research, 301: 19–35. https://doi.org/10.1016/j.precamres.2017.08.023 |
| ○ |
Aharon, P.; Schidlowski, M. & Singh, I.B. (1987). Chronostratigraphic markers in the end Precambrian carbon isotope record of the Lesser Himalaya. Nature, 327: 699–702. https://doi.org/10.1038/327699a0 |
| ○ |
Aitchison, J.C.; Suzuki, N.; Caridroit, M.; Danelian, T. & Noble, P. (2017). Paleozoic radiolarian biostratigraphy. In: Catalogue of Paleozoic radiolarian genera (Danelian, T. et al., Eds.). Geodiversitas, 39: 503–531. https://doi.org/10.5252/g2017n3a5 |
| ○ |
Álvaro, J.J. & Blanc-Valleron, M.M. (2002). Stratigraphic and structural framework of the Neoproterozoic Paracuellos Group, Iberian Chains, NE Spain. Bulletin de la Societé géologique de France, 173: 27–35. https://doi.org/10.2113/173.3.219 |
| ○ |
A´lvaro, J.J.; Monceret, E.; Monceret, S.; Verraes, G. & Vizcaïno, D. (2010). Stratigraphic record and palaeogeographic context of the Cambrian Epoch 2 subtropical carbonate platforms and their basinal counterparts in SW Europe, West Gondwana. Bulletin of Geosciences, 85: 573–584. https://doi.org/10.3140/bull.geosci.1179 |
| ○ |
Álvaro, J.J.; Shields-Zhou, G.; A.; Ahlberg, P.; Jensen, S. & Palacios, T. (2016). Ediacaran-Cambrian phosphorites from the western margins of Gondwana and Baltica. Sedimentology, 63: 350–377. https://doi.org/10.1111/sed.12217 |
| ○ |
An, Z.; Jiang, G.; Tong, J.; Tian, L.; Ye, Q.; Song, H. & Song, H. (2015). Stratigraphic position of the Ediacaran Miaohe biota and its constrains on the age of the upper Doushantuo δ13C anomaly in the Yangtze Gorges area, South China. Precambrian Research, 271: 243–253. https://doi.org/10.1016/j.precamres.2015.10.007 |
| ○ |
Anderson, R.P.; Macdonald, F.A.; Jones, D.S.; McMahon, S. & Briggs, D.E.G. (2017). Doushantuo-type microfossils from latest Ediacaran phosphorites of northern Mongolia. Geology, 45: 1079–1082. https://doi.org/10.1130/G39576.1 |
| ○ |
Anderson, R.P.; McMahon, S.; Macdonald, F.A.; Jones, D.S. & Briggs, D.E.G. (2019). Palaeobiology of latest Ediacaran phosphorites from the upper Khesen Formation, Khuvsgul Group, northern Mongolia. Journal of Systematic Palaeontology, 17: 501–532. https://doi.org/10.1080/14772019.2018.1443977 |
| ○ |
Ansari, A.H.; Pandey, S.K, Sharma, M.; Agrawal, S. & Kumar, Y. (2018). Carbon and oxygen isotope stratigraphy of the Ediacaran Bilara Group, Marwar Supergroup, India: Evidence for high amplitude carbon isotopic negative excursions. Precambrian Research, 308: 75–91. https://doi.org/10.1016/j.precamres.2018.02.002 |
| ○ |
Bhatt, D.K. & Mathur, A.K. (1990). Small shelly fossils of the Precambrian-Cambrian boundary beds from the Krol-Tal succession in the Nanital Syncline, Lesser Himalaya. Current Science, 59: 218–222. |
| ○ |
Brasier, M.; Cowie, J. & Taylor, M. (1994). Decision on the Precambrian-Cambrian boundary stratotype. Episodes, 17: 3–8. https://doi.org/10.18814/epiiugs/1994/v17i1.2/002 |
| ○ |
Brasier, M.D.; McIlroy, D.; Liu, A.G.; Antcliffe, J.B. & Menon, L.R. (2013). The oldest evidence of bioturbation on Earth: Comment. Geology, 41 (5), e289. https://doi.org/10.1130/G33606C.1 |
| ○ |
Braun, A.; Chen, J.; Waloszek, D. & Maas, A. (2007). First early Cambrian Radiolaria. In: The Rise and Fall of the Ediacaran Biota (Vickers-Rich, P. & Komarower, P.; eds.). Geological Society, London, Special Publications, 286: 143–149. https://doi.org/10.1144/SP286.10 |
| ○ |
Buatois, L.A.; Mángano, M.G.; Noffke, N. & Chafetz, H. (2011). The trace-fossil record of organism-matground interactions in space and time. SEPM Special Publication, 101: 15–28. https://doi.org/10.2110/sepmsp.101.015 |
| ○ |
Buatois, L.A.; Wisshak, M.; Wilson, M.A. & Mángano, M.G. (2017). Categories of architectural designs in trace fossils: a measure of ichnodisparity. Earth-Science Reviews, 164: 102–181. https://doi.org/10.1016/j.earscirev.2016.08.009 |
| ○ |
Buatois, L.A.; Almond, J.; Mángano, M.G.; Jensen, S. & Germs, G.J.B. (2018). Sediment disturbance by Ediacaran bulldozers and the roots of the Cambrian explosion. Scientific Reports, 8: 4514. https://doi.org/10.1038/s41598-018-22859-9 |
| ○ |
Budd, G.E. & Jensen, S. (2003). The limitations of the fossil record and the dating of the origin of the Bilateria. Systematics Association Special Volume, 66: 166–189.
|
| ○ |
Butterfield, N.J. (2003). Exceptional fossil preservation and the Cambrian Explosion. Integrative and Comparative Biology, 43: 166–177. https://doi.org/10.1093/icb/43.1.166 |
| ○ |
Butterfield, N.J. (2007). Macroevolution and macroecology through deep time. Palaeontology, 50: 41–55. https://doi.org/10.1111/j.1475-4983.2006.00613.x |
| ○ |
Butterfield, N.J. & Harvey, T.H.P. (2012). Small carbonaceous fossils (SCFs): A new measure of early Paleozoic paleobiology. Geology, 40: 71–74 https://doi.org/10.1130/G32580.1 |
| ○ |
Cai, Y.; Xiao, S.; Li, G. & Hua, H. (2019). Diverse biomineralizing animals in the terminal Ediacaran Period herald the Cambrian explosion. Geology, 47: 380–384. https://doi.org/10.1130/G45949.1 |
| ○ |
Cao, W.; Feng, Q.; Feng, F. & Ling, W. (2014). Radiolarian Kalimnasphaera from the Cambrian Shuijinguo Formation in South China. Marine Micropaleontology, 110: 3–7. https://doi.org/10.1016/j.marmicro.2013.06.005 |
| ○ |
Carbone, C. & Narbonne, G.M. (2014). When life got smart: the evolution of behavioral complexity through the Ediacaran and Early Cambrian of NW Canada. Journal of Paleontology, 88: 309–330. https://doi.org/10.1666/13-066 |
| ○ |
Caridroit, M.; Danelian, T.; O’Dogherty, L.; Cuvelier, J.; Aitchison, J.C.; Pouille, L.; Noble, P.; Dumitrica, P.; Suzuki, N.; Kuwahara, K.; Maletz, J. & Feng, Q. (2017). An illustrated catalogue and revised classification of Paleozoic radiolarian genera. In: Catalogue of Paleozoic radiolarian genera (Danelian, T. et al., Eds.). Geodiversitas, 39: 363–417. https://doi.org/10.5252/g2017n3a3 |
| ○ |
Chang, S.; Feng, Q. & Zhang, L. (2018). New siliceous microfossils from the Terreneuvian Yanjiahe Formation, South China: the possible earliest radiolarian fossil record. Journal of Earth Science, 29: 912–919. https://doi.org/10.1007/s12583-017-0960-0 |
| ○ |
Chauhan, D.S.; Morari Krishna & Ram Narayan (2001). Geological nature of the Pokaran Boulder bed; Palaeo-environmental, Palaeoclimatic and stratigraphic implication. Journal of the Geological Society of India, 58: 425–433.
|
| ○ |
Chen, M. & Liu, K. (1986). The geological significance of newly discovered microfossils from the upper Sinian (Doushantuo age) phosphorites. Scientia Geologica Sinica, 1: 46–53.
|
| ○ |
Chen, Z.; Zhou, C.M.; Meyer, M.; Xiang, K.; Schiffbauer, J.D.; Yuan, X.L. & Xiao, S.H. (2013). Trace fossil evidence for Ediacaran bilaterian animals with complex behaviors. Precambrian Research, 224: 690–701. https://doi.org/10.1016/j.precamres.2012.11.004 |
| ○ |
Chen, D.; Zhou, X.; Fu, Y.; Wang J. & Yan, D. (2015). New U-Pb zircon ages of the Ediacaran-Cambrian boundary strata in South China. Terra Nova, 27: 62–68. https://doi.org/10.1111/ter.12134 |
| ○ |
Cheng, M.; Li, C.; Zhou, L.; Feng, L.; Algeo, T.J.; Zhang, F.; Romaniello, S.; Jin, C.; Ling, H. & Jiang, S. (2017). Transient deep-water oxygenation in the early Cambrian Nanhua Basin, South China. Geochimica et Cosmochimica Acta, 210: 42–58. https://doi.org/10.1016/j.gca.2017.04.032 |
| ○ |
Chen Zheng-peng, Zhao Yuan-long, Luo Xiu-chun & Yang Guang-fu (2018). Trilobite Redlichia from the Cambrian “Tsinghsutung Formation” at Balang area in Jianhe, Guizhou and its biostratigraphic significance. Journal of Stratigraphy, 42: 352–359.
|
| ○ |
Condon, D.; Zhu, M.; Bowring, S.; Wang, W.; Yang, A. & Jin, Y. (2005). U-Pb Ages from the Neoproterozoic Doushantuo Formation, China. Science, 308 (5718): 95–98. https://doi.org/10.1126/science.1107765 |
| ○ |
Corsetti, F.A. & Hagadorn, J.W. (2003). The Precambrian-Cambrian transition in the southern Great Basin, USA. The Sedimentary Record, 1: 4–8. https://doi.org/10.2110/sedred.2003.1.4 |
| ○ |
Crimes, T.P. (1974). Colonisation of the early ocean floor. Nature, 248: 328–330. https://doi.org/10.1038/248328a0 |
| ○ |
Crimes, T.P. (1987). Trace fossils and correlation of late Precambrian and early Cambrian strata. Geological Magazine, 124:
97–119. https://doi.org/10.1017/S0016756800015922 |
| ○ |
Crimes, T.P. (1992a). Changes in the trace fossil biota across the Proterozoic-Phanerozoic boundary. Journal of the Geological Society, 149: 637–646. https://doi.org/10.1144/gsjgs.149.4.0637 |
| ○ |
Crimes, T.P. (1992b). The record of trace fossils across the Proterozoic-Cambrian boundary. In: Origin and early evolution of the Metazoa (Lipps, J.H. & Signor, P.H.; Eds.), Springer, Boston, MA, 177–202. https://doi.org/10.1007/978-1-4899-2427-8_6 |
| ○ |
Crimes, T.P.; Legg, L, Marcos, A. & Arboleya, M. (1977). ?Late Precambrian-low Lower Cambrian trace fossiIs from Spain. In: Trace Fossils 2 (Crimes, T.P. & & Harper, J.C.; Eds.). Geological Journal, Special Issue, 9: 91–138.
|
| ○ |
Danelian, T. & Morreira, D. (2004). Palaeontological and molecular arguments for the origin of silica secreting marine organisms. Comptes Rendus Palevol, 3: 229–236. https://doi.org/10.1016/j.crpv.2004.01.005 |
| ○ |
Darroch, S.A.F.; Sperling, E.A.; Boag, T.H.; Racicot, R.A.; Mason, S.J.; Morgan, A.S.; Tweedt, S.; Myrow, P.; Johnston, D.T.; Erwin, D.H. & Laflamme, M. (2015). Biotic replacement and mass extinction of the Ediacara biota. Proceedings of the Royal Society B-Biological Sciences, 282: 129–138. https://doi.org/10.1098/rspb.2015.1003 |
| ○ |
Darroch, S.A.F.; Boag, T.H.; Racicot, R.A.; Tweedt, S.; Mason, S.J.; Erwin, D.H. & Laflamme, M. (2016). A mixed Ediacaran-metazoan assemblage from the Zaris Sub-basin, Namibia. Palaeogeography, Palaeoclimatology, Palaeoecology, 459: 198–208. https://doi.org/10.1016/j.palaeo.2016.07.003 |
| ○ |
Dias da Silva, Í.; Díez Fernández, R.; Díez-Montes, A.; Gonzaález Clavijo, E. & Foster, D.A. (2016). Magmatic evolution in the N-Gondwana margin related to the opening of the Rheic Ocean- evidence from the Upper Parautochthon of the Galicia-Trás-os-Montes Zone and from the Central Iberian Zone (NW Iberian Massif). International Journal of Earth Sciences, 105: 1127–1151. https://doi.org/10.1007/s00531-015-1232-9 |
| ○ |
Dong, L.; Xiao, S.; Shen, B. & Zhou C. (2008). Silicified Horodyskia and Palaeopascichnus from upper Ediacaran cherts in South China: tentative phylogenetic interpretation and implications for evolutionary stasis. Journal of the Geological Society, London, 165: 367–378. https://doi.org/10.1144/0016-76492007-074 |
| ○ |
Dong, L.; Song, W.; Xiao, S.; Yuan, X.; Chen, Z. & Zhou, C. (2012). Micro- and macrofossils from the Piyuancun Formation and their implications for the Ediacaran-Cambrian boundary in Southern Anhui. Journal of Stratigraphy, 36: 600-610.
|
| ○ |
Dumitrica, P.; Caridroit, M. & De Wever, P. (2000). Archaeospicularia, new Radiolarian order; a new step for the classification of the Lower Paleozoic Radiolaria. Comptes Rendus de l’Académie des Sciences Paris, Comptes Rendus Palevol, 330: 563–569.
https://doi.org/10.1016/S1251-8050(00)00168-3 |
| ○ |
Erwin, D.H.; Laflamme, M.; Tweedt, S.M.; Sperling, E.A.; Pisani, D. & Peterson, K.J. (2011). The Cambrian conundrum: Early divergence and later ecological success in the early history of animals. Science, 334: 1091–1097. https://doi.org/10.1126/science.1206375 |
| ○ |
Gámez Vintaned, J.; Liñán, E.; Navarro, D. & Zhuravlev, A. (2018). The oldest Cambrian skeletal fossils of Spain (Cadenas Ibéricas, Aragón). Geological Magazine, 155: 1465–1474 https://doi.org/10.1017/S0016756817000358 |
| ○ |
Gehling, J.G.; Jensen, S.; Droser, M.L.; Myrow, P.M. & Narbonne, G.M. (2001). Burrowing below the basal Cambrian GSSP, Fortune Head, Newfoundland. Geological Magazine, 138: 213–218. https://doi.org/10.1017/S001675680100509X |
| ○ |
Geyer, G. & Peel, J.S. (2011). The Henson Gletscher formation. North Greenland and itsbearing on the global Cambrian Series 2–3 boundary. Bulletin of Geosciences, 86: 465–534. https://doi.org/10.3140/bull.geosci.1252 |
| ○ |
Gevirtzman, D.A. & Mount, J.F. (1986). Paleoenvironments of an earliest Cambrian (Tommotian) shelly fauna in the southwestern Great Basin, USA. Journal of Sedimentary Petrology, 56: 412–421. https://doi.org/10.1306/212F8931-2B24-11D7-8648000102C1865D |
| ○ |
Golubkova, E.Y.; Raevskaya, E.G. & Kuznetsov, A.B. (2010). Lower Vendian microfossil assemblages of East Siberia: Significance for solving regional stratigraphic problems. Stratigraphy and Geological Correlation, 18: 353–375. https://doi.org/10.1134/S0869593810040015 |
| ○ |
Golubkova, E.Y.; Zaitseva, T.S.; Kuznetsov, A.B.; Dovzhikova, E.G. & Maslov, A.V. (2015). Microfossils and Rb-Sr age of glauconite in the key section of the Upper Proterozoic of the northeastern part of the Russian plate (Keltmen-1 borehole). Doklady Earth Sciences, 462: 547–551. https://doi.org/10.1134/S1028334X15060045 |
| ○ |
Gonçalves, F. (1971). Subsídios para o conhecimento geológico do Nordeste Alentejano. Memória dos Serviços Geológicos de Portugal, 18: 1–62.
|
| ○ |
Gougeon, R.C.; Mángano, M.G.; Buatois, L.A.; Narbonne, G.M. & Laing, B.A. (2018a). Early Cambrian origin of the shelf sediment mixed layer. Nature Communications, 9: 1909. https://doi.org/10.1038/s41467-018-04311-8 |
| ○ |
Gougeon, R.; Néraudeau, D.; Dabard, M.P.; Pierson-Wickmann, A.C.; Polette, F.; Poujol, M. & Saint-Martin, J.P. (2018b). Trace fossils from the Brioverian (Ediacaran-Fortunian) in Brittany (NW France). Ichnos, 25: 11–24. https://doi.org/10.1080/10420940.2017.1308865 |
| ○ |
Gougeon, R.C.; Mángano, G.M.; Buatois, L.A.; Narbonne, G.M. & Laing, B.A. (2019). The role of Psammichnites in the origin of an Early Cambrian shelf sediment mixed layer. XV International Ichnofabric Workshop, Abstracts, 17–18. https://doi.org/10.1038/s41467-018-04311-8 |
| ○ |
Gradstein, F.M.; Ogg, J.G.; Schmitz, M., & Ogg, G. (Eds.). (2012). The geologic time scale 2012. Elsevier. 1144 pp. |
| ○ |
Grey, K. (2005). Ediacaran palynology of Australia. Memoirs of the Association of Australasian Palaeontologists, 31: 1–439. |
| ○ |
Grotzinger, J.P.; Fike, D.A. & Fischer, W.W. (2011). Enigmatic origin of the largest-known carbon isotope excursion in Earth’s history. Nature Geoscience, 4: 285–292. https://doi.org/10.1038/ngeo1138 |
| ○ |
Guilbaud, R.; Slater, B.J.; Poulton, S.W.; Harvey, T.H.P.; Brocks, J.J.; Nettersheim, B.J. & Butterfield, N.J. (2018). Oxygen minimum zones in the early Cambrian ocean. Geochemical Perspectives Letters, 6: 33–38. https://doi.org/10.7185/geochemlet.1806 |
| ○ |
Han, J.; Shu, D.; Zhang, Z. & Liu, J. (2004). The earliest-known ancestors of recent Priapulomorpha from the Early Cambrian Chengjiang Lagerstätte. Chinese Science Bulletin, 49: 1860–1868. https://doi.org/10.1007/BF03183414 |
| ○ |
Harvey, T.H.P.; Ortega-Hernández, J.; Lin, J.P.; Yuanlong, Z. & Butterfield, N.J. (2012). Burgess Shale-type microfossils from the middle Cambrian Kaili Formation, Guizhou Province, China. Acta Palaeontologica Polonica, 57: 423–436. https://doi.org/10.4202/app.2011.0028 |
| ○ |
Hawkins, A.D.; Xiao, S.; Jiang, G.; Wang, X. & Shi, X. (2017). New biostratigraphic and chemostratigraphic data from the Ediacaran Doushantuo Formation in intra-shelf and upper slope facies of the Yangtze platform: Implications for biozonation of acanthomorphic acritarchs in South China. Precambrian Research, 300: 28–39. https://doi.org/10.1016/j.precamres.2017.08.004 |
| ○ |
Hegenberger, W. (1993) Stratigraphy and sedimentology of the late Precambrian Witvlei and Nama Groups, east of Windhoek. Geological Survey of Namibia, Memoir, 17: 82.
|
| ○ |
Hofmann, H.J. & Patel, I.M. (1989). Trace fossils from the type ‘Etcheminian Series’ (Lower Cambrian Ratcliffe Brook Formation), Saint John area, New Brunswick, Canada. Geological Magazine, 126: 139–157. https://doi.org/10.1017/S0016756800006294 |
| ○ |
Högström, A.E.S.; Jensen, S.; Palacios, T. & Ebbestad, J.O.R. (2013). New information on the Ediacaran-Cambrian transition in the Vestertana Group, Finnmark, northern Norway, from trace fossils and organic-walled microfossils. Norwegian Journal of Geology, 93: 95–106.
|
| ○ |
Hughes, N.C. (2016). The Cambrian palaeontological record of the Indian subcontinent. Earth-Science Reviews, 159: 428–461.
https://doi.org/10.1016/j.earscirev.2016.06.004 |
| ○ |
Jensen, S. (2003). The Proterozoic and earliest Cambrian trace fossil record; patterns, problems and perspectives. Integrative and Comparative Biology, 43: 219–228. https://doi.org/10.1093/icb/43.1.219 |
| ○ |
Jensen, S.; Saylor, B.Z.; Gehling, J.G. & Germs, G.J.B. (2000). Complex trace fossils from the terminal Proterozoic of Namibia. Geology, 28: 143–146. https://doi.org/10.1130/0091-7613(2000)028<0143:CTFFTT>2.3.CO;2 |
| ○ |
Jensen, S. & Runnegar, B. N. (2005). A complex trace fossil from thee Spitskop Member (terminal Ediacaran-? Lower Cambrian) of southern Namibia. Geological Magazine, 142(5): 561–569. https://doi.org/10.1017/S0016756805000853 |
| ○ |
Jensen, S.; Droser, M.L. & Gehling, J.G. (2006). A critical look at the Ediacaran trace fossil record. In: Neoproterozoic Geobiology and Paleobiology (Xiao, S. & Kaufman, A.J., Eds.), Springer, 115–157. https://doi.org/10.1007/1-4020-5202-2_5 |
| ○ |
Jiang, G.; Shi, X.; Zhang, S.; Wang, Y. & Xiao, S. (2011). Stratigraphy and paleogeography of the Ediacaran Doushantuo Formation (ca. 635–551 Ma) in South China. Gondwana Research, 19: 831–849. https://doi.org/10.1016/j.gr.2011.01.006 |
| ○ |
Joshi, H. & Tiwari, M. (2016). Tianzhushania spinosa and other large acanthomorphic acritarchs of Ediacaran Period from the Infrakrol Formation, Lesser Himalaya, India. Precambrian Research, 286: 325–336. https://doi.org/10.1016/j.precamres.2016.09.024 |
| ○ |
Knoll, A.H. (1992). Vendian microfossils in metasedimentary cherts of the Scotia group, Prins Karls Forland, Svalbard. Palaeontology, 35: 751–774.
|
| ○ |
Korovnikov, I.V.; Sennikov, N.V.; Danelian T.; Obut O.T. & Pouille L. (2013). The biostratigraphic and palaeoenvironmental significance of lower Cambrian (Bortoman) trilobites from the Ak-Kaya section of the Altai Mountains (southern Siberia, Russia). Annales de Paléontologie, 99: 79–89. https://doi.org/10.1016/j.annpal.2012.12.001 |
| ○ |
Kumar, S. & Pandey, S.K. (2009). Note on occurrence of Arumberia banksi and associated fossils from the Jodhpur Sandstone, Marwar Supergroup, Western Rajasthan Journal of the Palaeontological Society of India, 54: 171–178.
|
| ○ |
Lehane, J.R. & Ekdale, A.A. (2016). Morphometric analysis of graphoglyptid trace fossils in two dimensions: implications for behavioral evolution in the deep sea. Paleobiology, 42: 317–334. https://doi.org/10.1017/pab.2015.52 |
| ○ |
Lenton, T.M.; Boyle, R.A.; Poulton, S.W.; Shields-Zhou, G.A. & Butterfield, N.J. (2014). Co-evolution of eukaryotes and ocean oxygenation in the Neoproterozoic era. Nature Geoscience, 7: 257–265. https://doi.org/10.1038/ngeo2108 |
| ○ |
Li, C.; Cheng, M.; Zhu, M. & Lyons, T.W. (2018). Heterogeneous and dynamic marine shelf oxygenation and coupled early animal evolution. Emerging Topics in Life Sciences, 2: 279–288. https://doi.org/10.1042/ETLS20170157 |
| ○ |
Linnemann, U.; Pereira, M.F.; Jeffries, T.; Drost, K. & Gerdes, A. (2008). Cadomian Orogeny and the opening of the Rheic Ocean: new insights in the diacrony of geotectonic processes constrained by LA-ICP-MS U-Pb zircon dating (Ossa-Morena and Saxo-Thuringian Zones, Iberian and Bohemian Massifs). Tectonophysics, 461: 21–43. https://doi.org/10.1016/j.tecto.2008.05.002 |
| ○ |
Linnemann, U.; Pidal, A.P.; Hofmann, M.; Drost, K.; Quesada, C.; Gerdes, A.; Marko, L.; Gärtner, A.; Zieger, J.; Ulrich, J.; Krause, R.; Vickers-Rich, P. & Horak, J.J.I.J. (2018). A ~565 Ma old glaciation in the Ediacaran of peri Gondwanan West Africa. International Journal of Earth Sciences, 107: 885–911. https://doi.org/10.1007/s00531-017-1520-7 |
| ○ |
Linnemann, U.; Ovtcharova, M.; Schaltegger, U.; Gartner, A.; Hautmann, M.; Geyer, G.; Vickers-Rich, P.; Rich, T.; Plessen, B.; Hofmann, M.; Zieger, J.; Krause, R.; Kriesfeld, L. & Smith, J. (2019). New high-resolution age data from the Ediacaran-Cambrian boundary indicate rapid, ecologically driven onset of the Cambrian explosion. Terra Nova, 31: 49–58. https://doi.org/10.1111/ter.12368 |
| ○ |
Liñan, E. & Tejero, R. (1988). Las formaciones precámbricas del antiforme de Paracuellos (Cadenas ibéricas). Boletín de la Real Sociedad Española de Historia Natural (Sección Geológica), 84: 39–49.
|
| ○ |
Liu, P. & Moczydłowska, M. (2019). Ediacaran microfossils from the Doushantuo Formation chert nodules in the Yangtze Gorges area, South China, and new biozones. Fossils and Strata, 65: 1–172. https://doi.org/10.1002/9781119564225 |
| ○ |
Liu, P.; Chen, S.; Zhu, M.; Li, M.; Yin, C. & Shang, X. (2014a). High-resolution biostratigraphic and chemostratigraphic data from the Chenjiayuanzi section of the Doushantuo Formation in the Yangtze Gorges area, South China: Implication for subdivision and global correlation of the Ediacaran System. Precambrian Research, 249: 199–214. https://doi.org/10.1016/j.precamres.2014.05.014 |
| ○ |
Liu, P.; Xiao, S.; Yin, C.; Chen, S.; Zhou, C. & Li, M. (2014b). Ediacaran acanthomorphic acritarchs and other microfossils from chert nodules of the upper Doushantuo Formation in the Yangtze Gorges area, South China. Journal of Paleontology, 88
(sp72): 1–139. https://doi.org/10.1666/13-009 |
| ○ |
López Sopeña F.; Villarino, G.M.; Del Olmo, A. et al. (1991). Mapa Geológico de España a escala 1:50.000 (2ª Serie). Hoja 753 - Miajadas. IGME, Madrid.
|
| ○ |
Lotze, F. (1961). Das Kambrium Spaniens, Teil I: Stratigraphie. Akademie der Wissenschaften. und der Literatur Mainz, Abhandlungen der mathematisch-naturwissenschaftlichen Klasse, 6: 283–498.
|
| ○ |
Luo, C. & Reitner, J. (2019). Three-dimensional preserved stem-group hexactinellid sponge fossils from lower Cambrian (Stage 2) phosphorites of China. Paläontologische Zeitschrift, 93: 187–194 https://doi.org/10.1007/s12542-018-00441-y |
| ○ |
Ma, Q.; Feng, Q.; Cao, W.; Zhang, W.; Zhang L.; Ye.Y. & Gu, S. (2019). Radiolarian fauna from the Chiungchussuan Shuijingtuo Formation (Cambrian Series 2) in Western Hubei Province, South China. Science China Earth Sciences, 62, https://doi.org/10.1007/s11430-018-9335-0. |
| ○ |
Maheshwari, A.; Sial, A.N. & Mathur. S.C. (2003). Carbonate and Oxygen Isotope profiles from the terminal Precambrian Marwar Supergroup, Rajasthan. India. International Journal of Carbonates and Evaporates, 4: 18: 64–73. https://doi.org/10.1007/BF03178383 |
| ○ |
Maletz, J. (2011). Radiolarian skeletal structures and biostratigraphy in the early Palaeozoic (Cambrian-Ordovician). Palaeoworld, 20: 116–133. https://doi.org/10.1016/j.palwor.2010.12.007 |
| ○ |
Maletz, J. (2015). The earliest radiolarians: The race is still open. Proceedings of 14th INTERRAD, March 22–26 2015, Antalya, Turkey: 62–63.
|
| ○ |
Maletz, J. (2017). The identification of putative Lower Cambrian Radiolaria. Revue de Micropaléontologie, 60: 233–240. https://doi.org/10.1016/j.revmic.2017.04.001 |
| ○ |
Mángano, M.G. & Buatois, L.A. (2014). Decoupling of body-plan diversification and ecological structuring during the Ediacaran-Cambrian transition: evolutionary and geobiological feedbacks. Proceedings of the Royal Society B, 281: 20140038. https://doi.org/10.1098/rspb.2014.0038 |
| ○ |
Mao, Y.Q. (2017). Study on Cambrian system Kaili formation Brachiopoda in Southeast of Guizhou Province. PhD, Jiling University. |
| ○ |
Mao, Y.Q.; Zhao, Y.L.; Wang C.W. & Topper, T. (2017). A fresh look at Nisusia Walcott, 1905 from the Cambrian Kaili Formation in Guizhou. Palaeoworld, 26:12–24. https://doi.org/10.1016/j.palwor.2016.03.001 |
| ○ |
Marusin, V.V. (2011). Evolution of organism-sediment interaction in transition to Phanerozoic biosphere. In: Neoproterozoic sedimentary basins: stratigraphy, geodynamics and petroleum potential. Proceedings of the International Conference, Novosibirsk, 61–62.
|
| ○ |
Marshall, C.R. (2006). Explaining the Cambrian “Explosion” of Animals. Annual Review of Earth & Planetary Sciences, 34: 355–384.
https://doi.org/10.1146/annurev.earth.33.031504.103001 |
| ○ |
Matas, J.; Moreno, F.; Leyva, F. et al. (2005). Mapa Geológico de España a escala 1:50.000 (2ª Serie). Hoja 755 - Navalvillar de Pela. IGME, Madrid.
|
| ○ |
Mathur, S.C.; Parihar, V.S.; Nama, S.L.; Soni, A.; Ram, H. & Mathur Saurabh (2017). The global Ediacaran Event in the geological record of the Jodhpur Group of the Marwar Supergroup: Enigmas in geology’s prelude to the Cambrian Explosion. Keynote address, Proceedings of the Seminar on Recent Updates in Precambrian Geology, Udaipur, India, 5–9.
|
| ○ |
McDougall, N.; Brenchley, P.J.; Rebelo, J.A. & Romano, M. (1987). Fans and fan deltas- precursors to the Armorican Quartzite (Ordovician) in western Iberia. Geological Magazine, 124: 347–359. https://doi.org/10.1017/S0016756800016678 |
| ○ |
McIlroy, D. & Brasier, M.D. (2017). Ichnological evidence for the Cambrian explosion in the Ediacaran to Cambrian succession of Tanafjord, Finnmark, northern Norway. Geological Society of London Special Publications, 448(1): 351–368. |
| ○ |
Miller, M.F. & Smail, S.E. (1997). A semiquantitative field method for evaluating bioturbation on bedding planes. Palaios, 12: 391–396. https://doi.org/10.2307/3515338 |
| ○ |
Moczydłowska, M. & Nagovitsin, K.E. (2012). Ediacaran radiation of organic-walled microbiota recorded in the Ura Formation, Patom Uplift, East Siberia. Precambrian Research, 198–199: 1–24. https://doi.org/10.1016/j.precamres.2011.12.010 |
| ○ |
Moczydlowska, M.; Vidal, G. & Rudavskaya, V.A. (1993). Neoproterozoic (Vendian) phytoplankton from the Siberian Platform, Yakutia. Palaeontology, 36: 495–521.
|
| ○ |
Müller, K.J. & Hinz, I. (1991). Upper Cambrian conodonts from Sweden. Fossils and Strata, 28: 1–153 |
| ○ |
Muscente, A.D.; Hawkins, A.D. & Xiao, S. (2015). Fossil preservation through phosphatization and silicification in the Ediacaran Doushantuo Formation (South China): A comparative synthesis. Palaeogeography, Palaeoclimatology, Palaeoecology, 434: 46–62.
https://doi.org/10.1016/j.palaeo.2014.10.013 |
| ○ |
Narbonne, G.M.; Xiao, S.; Shields, G.A. & Gehling, J.G. (2012). The Ediacaran Period. In: The Geologic Time Scale (Gradstein, F.M.; Ogg, J.G.; Schmitz, M.D. & Ogg, G.M., Eds.). Elsevier, Boston, 413–435. https://doi.org/10.1016/B978-0-444-59425-9.00018-4 |
| ○ |
Néraudeau, D.; Dabard, M.P.; El Albani, A.; Gougeon, R.; Mazurier, A.; Pierson-Wickmann, A.C.; Poujol, M.; Saint Martin, J.P. & Saint Martin, S. (2018). First evidence of Ediacaran-Fortunian elliptical body fossils in the Brioverian series of Brittany, NW France. Lethaia, 51: 513–522. https://doi.org/10.1111/let.12270 |
| ○ |
Noble, P.; Aitchison, J.C.; Danelian, T.; Dumitrica, P.; Maletz, J.; Suzuki, N.; Caridroit, M.; O’Dogherty, L. & Cuvelier, J. (2017). Taxonomy of Paleozoic radiolarian genera. In: Catalogue of Paleozoic radiolarian genera (Danelian, T. et al., Eds.). Geodiversitas, 39: 419–502. https://doi.org/10.5252/g2017n3a4 |
| ○ |
Obut, O.T. & Iwata, K. (2000). Lower Cambrian Radiolaria from the Gorny Altai (southern West Siberia). News of Paleontology and Stratigraphy, 2–3: 33–38.
|
| ○ |
Oji, T.; Dornbos, S.Q.; Yada, K.; Hasegawa, H.; Gonchigdorj, S.; Mochizuki, T.; Takayanagi, H. & Iryu, Y. (2018). Penetrative trace fossils from the late Ediacaran of Mongolia: early onset of the agronomic revolution. Royal Society Open Science, 5:
172250. https://doi.org/10.1098/rsos.172250 |
| ○ |
Ortega Gironés, E. & González Lodeiro, F. (1986). La discordancia intra-Alcudiense en el dominio meridional de la Zona Centroibérica. Geológica Astúrica, 27: 27–32.
|
| ○ |
Ouyang, Q.; Guan, C.; Zhou, C. & Xiao, S. (2017). Acanthomorphic acritarchs of the Doushantuo Formation from an upper slope section in northwestern Hunan Province, South China, with implications for early-middle Ediacaran biostratigraphy. Precambrian Research, 298: 512–529. https://doi.org/10.1016/j.precamres.2017.07.005 |
| ○ |
Palero, F.J. (1993). Tectónica pre-hercínica de las series infraordovícicas del anticlinal de Alcudia y la discordancia intraprecámbrica en su pate oriental (-sector meridional de la Zona Centroibérica). Boletín Geológico y Minero, 104: 227–242.
|
| ○ |
Pareek, H.S. (1981). Configuration and sedimentary stratigraphy of western Rajasthan. Journal of the Geological Society of India, 22: 517–523.
|
| ○ |
Pareek, H.S. (1984). Pre-Quarternary geology and mineral resources of north- western Rajasthan. Memoirs of the Geological Survey of India, 115: 1–99.
|
| ○ |
Parihar, V.S.; Gour, V.; Nama, S.L. & Mathur, S.C. (2015). New report of Arumberia Banksi (Ediacaran affinity) mat structures from Girbhakar sandstone of the Marwar Supergroup, Bhopalgarh area, Jodhpur, western Rajasthan, India. In: Frontiers of Earth -Science. Indian Science Congress (Shrivastava, K.L. & Shrivastava, P.K., Eds.), 385–392.
|
| ○ |
Parry, L.A.; Boggiani, P.C.; Condon, D.J.; Garwood, R.J.; Leme, J.D.; McIlroy, D.; Brasier, M.D.; Trindade, R.; Campanha, G.A.C.; Pacheco, M.L.A.F.; Diniz, C.Q.C. & Liu, A.G. (2017). Ichnological evidence for meiofaunal bilaterians from the terminal Ediacaran and earliest Cambrian of Brazil. Nature Ecology & Evolution, 1: 1455–1464. https://doi.org/10.1038/s41559-017-0301-9 |
| ○ |
Peng S.C. (2000). A new chronosratigraphic subdivision of Cambrian for China. Miscelánea, 154: 260–270. |
| ○ |
Peng, S.C. & Zhao, Y.L. (2018). The proposed global standard stratotype - section and point (GSSP) for the conterminous base of Miaolingian series and Wuliuan stage at balang, Jianhe, Guizhou, China was ratified by IUGS. Journal of Stratigraphy, 42:
325–327.
|
| ○ |
Peng Shan-chi, Babcock L E, Zhu Xue-jian, Lei Qian-ping & Dai Tao (2017). Revision of the oryctocephalid trilobite genera Arthricocephalus Bergeron and Oryctocarella Tomashpolskaya and Karpinski (Cambrian) from South China and Siberia. Journal of Paleontology, 91: 933–959. https://doi.org/10.1017/jpa.2017.44 |
| ○ |
Pereira, M.F. & Silva, J.B. (2001). The Northeast Alentejo Neoproterozoic-Lower Cambrian succession (Portugal): implications for regional correlations in the Ossa-Morena Zone (Iberian Massif). Sociedad Geologica de Espana. Geogaceta, 30: 106–111.
|
| ○ |
Pereira, M.F.; Chichorro, M.; Linnemann, U.; Eguíluz, L. & Silva, J.B. (2006). Inherited arc signature in Ediacaran and Early Cambrian basins of the Ossa-Morena Zone (Iberian Massif, Portugal): paleogeographic link with European and North African Cadomian correlatives. Precambrian Research, 144: 297–315. https://doi.org/10.1016/j.precamres.2005.11.011 |
| ○ |
Pereira, M.F.; Chichorro, M, Solá, A.R.; Silva, J.B.; Sánchez-García, T. & Bellido, F. (2011). Tracing the Cadomian magmatism with detrital/inherited zircon ages by in-situ U-Pb SHRIMP geochronology (Ossa-Morena Zone, SW Iberian Massif). Lithos, 123: 204–217. https://doi.org/10.1016/j.lithos.2010.11.008 |
| ○ |
Pereira, M.F.; Linnemann, U.; Hofmann, M.; Chichorro, M.; Sola, A.R.; Medina, J. & Silva, J.B. (2012). The provenance of Late Ediacaran and Early Ordovician siliciclastic rocks in the Southwest Central Iberian Zone: Constraints from detrital zircon data on northern Gondwana margin evolution during the late Neoproterozoic. Precambrian Research, 192–195: 166–189. https://doi.org/10.1016/j.precamres.2011.10.019 |
| ○ |
Pereira, M.F.; Castro, A.; Fernández, C. & Rodríguez, C. (2018). Multiple Paleozoic magmatic-orogenic events in the Central Extremadura batholith (Iberian Variscan belt, Spain). Journal of Iberian Geology, 44: 309–333. https://doi.org/10.1007/s41513-018-0063-5 |
| ○ |
Pickerill, R.K. (1980). Phanerozoic flysch trace fossil diversity - observations based on an Ordovician flysch ichnofauna from the Aroostook-Matapedia Carbonate Belt of northern New Brunswick. Canadian Journal of Earth Sciences, 17: 1259–1270.
https://doi.org/10.1139/e80-131 |
| ○ |
Pouille, L.; Obut, O.; Danelian, T. & Sennikov, N. (2011). Lower Cambrian (Botomian) polycystine Radiolaria from the Altai Mountains (southern Siberia, Russia). Comptes Rendus Palevol, 10: 627–633. https://doi.org/10.1016/j.crpv.2011.05.004 |
| ○ |
Prasad, B. & Asher, R. (2016). Record of Ediacaran complex acanthomorphic acritarchs from the lower Vindhyan succession of the Chambal valley (east Rajasthan), India and their biostratigraphic significance. Journal of the Palaeontological Society of India, 61 (1): 29–62.
|
| ○ |
Prave, A.R.; Hoffmann, K.H.; Hegenberger, W. & Fallick A.E. (2011). The Witvlei Group of East-Central Namibia. In: The Geological Record of Neoproterozoic Glaciations (Arnaud, E.; Halverson, G.P. & Shields-Zhou G., Eds.). Geological Society, London, Memoirs, 36: 211–216. https://doi.org/10.1144/M36.15 |
| ○ |
Reitner, J.; Luo, C. & Duda, J.P. (2012): Early sponge remains from the Neoproterozoic-Cambrian Phosphate deposits of the Fontanarejo area (Central Spain). Journal of Guizhou University (Natural Science), 29 (supplement 1): 184–186
|
| ○ |
Rhoads, D.C. & Young, D.K. (1970). Influence of deposit-feeding organisms on sediment stability and community trophic structure. Journal of Marine Research, 28: 150–178.
|
| ○ |
Robson, S.P.; Nowlan, G.S. & Pratt, B.R. (2003). Middle to Upper Cambrian linguliformean brachiopods from the Deadwood Formation of subsurface Alberta and Saskatchewan, Canada. Journal of Paleontology, 77: 201–211. https://doi.org/10.1017/S0022336000043602 |
| ○ |
Rogov, V.; Marusin, V.; Bykova, N.; Goy, Y. & Nagovitsin, K. (2012). The oldest evidence of bioturbation on Earth. Geology, 40: 395–398. https://doi.org/10.1130/G32807.1 |
| ○ |
Rogov, V.I.; Karlova, G.A.; Marusin, V.V.; Kochnev, B.B.; Nagovitsin, K.E. & Grazhdankin, D.V. (2015). Duration of the first biozone in the Siberian hypostratotype of the Vendian. Russian Geology and Geophysics, 56: 573–583. https://doi.org/10.1016/j.rgg.2015.03.016 |
| ○ |
Rolim, F.G. & Theodorovicz, A. (2012). Geoparque Bodoquena-Pantanal (MS) - proposta. In: Schobbenhaus, C.; Silva, A.R. (Eds). Geoparques do Brasil - propostas. Rio de Janeiro, Brasil. Serviço Geológico do Brasil (CPRM), 2012: 223–282. |
| ○ |
Sánchez-García, T.; Pereira, M.F.; Bellido, F.; Chichorro, M.; Silva, J.B.; Valverde-Vaquero, P.; Pin, Ch. & Solá, A.R. (2013). Early Cambrian granitoids of North Gondwana margin in the transition from a convergent setting to intra-continental rifting (Ossa-Morena Zone, SW Iberia). International Journal of Earth Sciences, 103: 1203–1218. https://doi.org/10.1007/s00531-013-0939-8 |
| ○ |
Sarkar, S.; Bose, P.K.; Samanta, P.; Sengupta, P. & Eriksson, G. (2008). Microbial mediated structures in the Ediacaran Sonia sandstone, Rajasthan, India and their implications for Proterozoic sedimentation. Precambrian Research, 162: 248–263. https://doi.org/10.1016/j.precamres.2007.07.019 |
| ○ |
Schoenberg, R.; Zink, S.; Staubwasser, M. & von Blanckenburg F. (2008). The stable Cr isotope inventory of solid Earth reservoirs determined by double spike MC-ICPMS. Chemical Geology, 249: 294–306. https://doi.org/10.1016/j.chemgeo.2008.01.009 |
| ○ |
Seilacher, A. (1977). Evolution of trace fossil communities. In: Developments in Palaeontology and Stratigraphy (Hallam, A.; Ed.), Elsevier, 359–376. https://doi.org/10.1016/S0920-5446(08)70331-5 |
| ○ |
Sennikov, N.V.; Korovnikov, I.V.; Obut, O.T.; Tokarev, D.A.; Novozhilova, N.V. & Danelian, T. (2017). The Lower Cambrian oft he Salair and Gorny Altai (Siberia) revisited. In: Insights into the geology and paleontology of Siberia from French-Siberian collaboration in Earth Sciences (Danelian, T. et al., Eds.). Bulletin de la Société géologique de France, 188: 109–118. https://doi.org/10.1051/bsgf/2017002 |
| ○ |
Sharma, M. & Mathur, S.C. (2014). A Paleontological note on Arumberia like Mat of Ediacaran age of the Jodhpur Group of the Marwar Supergroup, W. Rajasthan, India. In: Georesources. Indian Science Congress (Shrivastava, K.L. & Arun Kumar eds.). Scientific Publications, Jodhpur, 1: 167–175.
|
| ○ |
Sierra, R.; Matz, M.V.; Aglyamova, G.; Pillet, L.; Decelle, J.; Not, F.; de Vargas, C. & Pawlowski, J. (2013). Deep relationships of Rhizaria revealed by phylogenomics: A farewell to Haeckel’s Radiolaria. Molecular Phylogenetics and Evolution, 67: 53–59.
https://doi.org/10.1016/j.ympev.2012.12.011 |
| ○ |
Sims, D.W.; Reynolds, A.M.; Humphries, N.E.; Southall, E.J.; Wearmouth, V.J.; Metcalfe, B. & Twichett, R.J. (2014). Hierarchical random walks in trace fossils and the origin of optimal search behaviour. Proceedings of the National Academy of Sciences USA, 111(30): 11073–11078. https://doi.org/10.1073/pnas.1405966111 |
| ○ |
Slater, B.J. & Willman, S. (2019). Early Cambrian small carbonaceous fossils (SCFs) from an impact crater in western Finland. Lethaia, https://doi.org/10.1111/let.12331. |
| ○ |
Slater, B.J.; Harvey, T.H.P.; Guilbaud, R. & Butterfield, N.J. (2017). A cryptic record of Burgess Shale-type diversity from the early Cambrian of Baltica. Palaeontology, 60: 117–140. https://doi.org/10.1111/pala.12273 |
| ○ |
Slater, B.J.; Harvey, T.H.P. & Butterfield, N.J. (2018a). Small carbonaceous fossils (SCFs) from the Terreneuvian (lower Cambrian) of Baltica. Palaeontology, 61: 417–439. https://doi.org/10.1111/pala.12350 |
| ○ |
Slater, B.J.; Willman, S.; Budd, G.E. & Peel, J.S. (2018b). Widespread preservation of small carbonaceous fossils (SCFs) in the early Cambrian of North Greenland. Geology, 46: 107–110. https://doi.org/10.1130/G39788.1 |
| ○ |
Smith, M.R.; Harvey, T.H. & Butterfield, N.J. (2015). The macro-and microfossil record of the Cambrian priapulid Ottoia. Palaeontology, 58: 705–721. https://doi.org/10.1111/pala.12168 |
| ○ |
Smith, E.F.; Nelson, L.L.; Strange, M.A.; Eyster, A.E.; Rowland, S.M.; Schrag, D.P. & Macdonald, F.A. (2016). The end of the Ediacaran: two new exceptionally preserved body fossil assemblages from Mount Dunfee, Nevada, USA. Geology, 44: 911–914. https://doi.org/10.1130/G38157.1 |
| ○ |
Spjeldnaes, N. (1963). A New Fossil (Papillomembrana sp.) from the Upper Precambrian of Norway. Nature, 200 (4901): 63–64.
https://doi.org/10.1038/200063a0 |
| ○ |
Streng, M. (1996). Erläuterungen zur geologischen Karte des Gebietes NE’ and SW’ Codos (Östliche Iberische Ketten, NE-Spanien). Diplomkartierung Paläontologisches Institut, Universität Würzburg.
|
| ○ |
Talavera, C.; Montero, P.; Martínez Poyatos, D. & Williams, I. S. (2012). Ediacaran to Lower Ordovician age for rocks ascribed to the Schist-Graywacke Complex (Iberian Massif, Spain): Evidence from detrital zircon SHRIMP U-Pb geochronology. Gondwana Research, 22: 928–942. https://doi.org/10.1016/j.gr.2012.03.008 |
| ○ |
Tarhan, L.G.; Droser, M.L.; Cole, D.B. & Gehling, J.G. (2018). Ecological expansion and extinction in the late Ediacaran: weighing the evidence for environmental and biotic drivers. Integrative and Comparative Biology, 58: 688–702. https://doi.org/10.1093/icb/icy020 |
| ○ |
Taylor, A.M. & Goldring, R. (1993). Description and analysis of bioturbation and ichnofabric. Journal of the Geological Society, 150: 141–148. https://doi.org/10.1144/gsjgs.150.1.0141 |
| ○ |
Teal, L.R.; Bulling, M.T.; Parker, E.R. & Solan, M. (2008). Global patterns of bioturbation intensity and mixed depth of marine soft sediments. Aquatic Biology, 2: 207–218. https://doi.org/10.3354/ab00052 |
| ○ |
Tena-Dávila Ruiz, M.; Corrétge, L. G.; González Lodeiro, F.; Martínez Catalán, J.R.; Granados, L.; Gil Gil, D. et al. (1980). Mapa Geológico de España a escala 1:50.000. Hoja 704 - Cáceres). IGME, Madrid.
|
| ○ |
Tiwari, M. & Knoll, A. (1994). Large acanthomorphic acritarchs from the Infrakrol Formation of the Lesser Himalaya and their stratigraphic significance. Journal of Himalayan Geology, 5: 193–201.
|
| ○ |
Uchman, A. (1995). Taxonomy and paleoecology of flysch trace fossils: The Marnoso-arenacea Formation and associated facies (Miocene, northern Apennines, Italy). Beringeria, 15: 1–115.
|
| ○ |
Vannier, J.; Calandra, I.; Gaillard, C. & Żylińska, A. (2010). Priapulid worms: pioneer horizontal burrowers at the Precambrian-Cambrian boundary. Geology, 38: 712–714. https://doi.org/10.1130/G30829.1 |
| ○ |
Vorob’eva, N.G.; Sergeev, V.N. & Knoll, A.H. (2009). Neoproterozoic microfossils from the margin of the East European Platform and the search for a biostratigraphic model of lower Ediacaran rocks. Precambrian Research, 173: 163–169. https://doi.org/10.1016/j.precamres.2009.04.001 |
| ○ |
Walde, D.H.G, Do Carmo, D. A.; Guimarães, E. M.; Vieira, L.C.; Erdtmann, B.D.; Sanchez, E.A.M, Adorno, R.R & Tobias, T.C.
(2015). New aspects of Neoproterozoic-Cambrian transition in the Corumbá region (state of Mato Grosso do Sul, Brazil). Annales de Paléontologie, 101: 213–224. https://doi.org/10.1016/j.annpal.2015.07.002 |
| ○ |
Witkosky, R. & Wernicke, B.P. (2018). Subsidence history of the Ediacaran Johnnie Formation and related strata of southwest Laurentia: Implications for the age and duration of the Shuram isotopic excursion and animal evolution. Geosphere, 14: 2245–2276.
https://doi.org/10.1130/GES01678.1 |
| ○ |
Won, M.Z. & Below, R. (1999). Cambrian radiolarians from the Georgina Basin, Queensland, Australia. Micropaleontology, 45:
325–363. https://doi.org/10.2307/1486119 |
| ○ |
Xiao, S.; Muscente, A.D.; Chen, L.; Zhou, C.; Schiffbauer, J.D.; Wood, A.D.; Polys, N.F. & Yuan, X. (2014a). The Weng’an biota and the Ediacaran radiation of multicellular eukaryotes. National Science Review, 1: 498–520. https://doi.org/10.1093/nsr/nwu061 |
| ○ |
Xiao, S.; Zhou, C.; Liu, P.; Wang, D. & Yuan, X. (2014b). Phosphatized acanthomorphic acritarchs and related microfossils from the Ediacaran Doushantuo Formation at Weng’an (South China) and their implications for biostratigraphic correlation. Journal of Paleontology, 88: 1–67. https://doi.org/10.1666/12-157R |
| ○ |
Xiao, S.; Narbonne, G.M.; Zhou, C.; Laflamme, M.; Grazhdankin, D.V.; Moczydlowska-Vidal, M. & Cui, H. (2016). Towards an Ediacaran time scale: problems, protocols, and prospects. Episodes, 39: 540–555. https://doi.org/10.18814/epiiugs/2016/v39i4/103886 |
| ○ |
Yuan, X.; Chen, Z.; Xiao, S.; Zhou, C. & Hua, H. (2011). An early Ediacaran assemblage of macroscopic and morphologically differentiated eukaryotes. Nature, 470: 390–393. https://doi.org/10.1038/nature09810 |
| ○ |
Zhang, K. & Feng, Q.L. (2019). Early Cambrian radiolarians and sponge spicules from the Niujiaohe Formation in South China. Palaeoworld (in press). https://doi.org/10.1016/j.palwor.2019.04.001 |
| ○ |
Zhang, Y.; Yin, L.; Xiao, S. & Knoll, A.H. (1998). Permineralized Fossils from the Terminal Proterozoic Doushantuo Formation, South China. Journal of Paleontology Memoirs, 72: 1–52. https://doi.org/10.1017/S0022336000059977 |
| ○ |
Zhao Yuan-long, Yu You-yi, Yuan Jin-liang, Yang Xing-lian, Guo Qing-jun (2001). Cambrian Stratigraphy at Huanglian, Guizhou Province, China: Referecec section for bases of the Naogaoan and Duyunian Stages. Palaeoworld, 13: 172–181.
|
| ○ |
Zhao, Y.L.; Yuan, J.L.; Esteve, J. et al. (2017). The oryctocephalid trilobite zonation across the Cambrian Series 2- Series 3 boundary at Balang, South China: a reappraisal. Lethaia, 50: 400–406. https://doi.org/10.1111/let.12227 |
| ○ |
Zhou, C.; Chen, Z. & Xue, Y. (2002). New microfossils from the late Neoproterozoic Doushantuo Formation at Chaoyang phosphorite deposit in Jiangxi Province, South China. Acta Palaeontologica Sinica, 41: 178–192.
|
| ○ |
Zhou, C.; Xie, G.; McFadden, K.A.; Xiao, S. & Yuan, X. (2007). The diversification and extinction of Doushantuo-Pertatataka acritarchs in South China: causes and biostratigraphic significance. Geological Journal, 42: 229–262. https://doi.org/10.1002/gj.1062 |
| ○ |
Zhou, C.; Xiao, S.; Wang, W.; Guan, C.; Ouyang, Q. & Chen, Z. (2017). The stratigraphic complexity of the middle Ediacaran carbon isotopic record in the Yangtze Gorges area, South China, and its implications for the age and chemostratigraphic significance of the Shuram excursion. Precambrian Research, 288: 23–38. https://doi.org/10.1016/j.precamres.2016.11.007 |
| ○ |
Zhou, C.; Yuan, X.; Xiao, S.; Chen, Z. & Hua, H. (2019). Ediacaran integrative stratigraphy and timescale of China. Science China Earth Sciences, 62: 7–24. https://doi.org/10.1007/s11430-017-9216-2 |
| ○ |
Zhu, M.; Zhuravlev, A.Y.; Wood, R.A.; Zhao, F. & Sukhov, S.S. (2017). A deep root for the Cambrian explosion: Implications of new bio- and chemostratigraphy from the Siberian Platform. Geology, 45: 459–462. https://doi.org/10.1130/G38865.1 |
| ○ |
Zybin, V.A.; Sennikov, N.V.; Iwata, K.; Obut, O.T.; Kurtigeshev, V.S.; Khlebnikova, T.V. & Sugai, Y. 2000. New data on the microfauna and geologic structure of the areas of development of the Lower Cambrian Shashkunar siliceous-carbonate-terrigenous Formation (Gorny Altai). Russian Geology and Geophysics, 41: 499–516.
|