First record of Triassic marine reptiles (Nothosauria, Sauropterygia) from the Alpujarride Complex (Internal Zones of the Betic Cordillera, Spain)
Primer registro de reptiles marinos triásicos (notosaurios, sauropterigios) del Complejo Alpujárride (Zonas Internas de la Cordillera Bética, España)
M. Reolid1,*, J. Reolid2
1 Departamento de Geología, Universidad de Jaén, Campus Las Lagunillas sn, 23071 Jaén, Spain. Email: mreolid@ujaen.es. ORCID ID: https://orcid.org/0000-0003-4211-3946
2 Departamento de Estratigrafía y Paleontología, Universidad de Granada, Campus Fuentenueva sn, 18071, Granada, Spain. Email: jreolid@ugr.es. ORCID ID: https://orcid.org/0000-0003-2329-4163
* Corresponding author
| |
ABSTRACT
This work reports the first record in the Internal Zones of the Betic Cordillera of a vertebrate fossil remain, more exactly a rib, corresponding to a marine reptile (Sauropterygia). The development of epicontinental platforms restricted to the Tethys Ocean during the fragmentation of Pangea since the late Lower Triassic favoured the radiation of Sauropterygia, diapsid marine reptiles including Placodontiformes, Pachypleurosauria, Nothosauroidea, and Pistosauroidea. The Triassic of the Alpujarride Complex, in the Internal Zones of the Betic Cordillera, SE Spain, comprises the carbonate deposits of these epicontinental platforms developed during the Middle-Late Triassic in the northern margin of the Mesomediterranean Massif. The studied stratigraphic interval is located at Sierra de Lújar (Granada Province) and consists of 17.5-m thick succession of alternating fossiliferous marls and limestones with local intense bioturbation and abundance of macroinvertebrates. The studied interval represents shallow facies of the inter- to subtidal environment as deduced by the record of Lofer cycles and the record of carbonated breccia at the top of the sequence likely related to storm events. Lofer cycles display lamination at their bases and intense bioturbation and abundant bivalves towards the top. A 15-cm long dorsal rib of a sauropterygian, likely a nothosaur, was recorded in this succession. It is well preserved, without evidence of original fragmentation, borings, or encrustations. This study shows for the first time fossil bones of marine reptiles in the Alpujarride Complex, which inhabited the shallow-water environments during the Triassic, equivalent to the marine reptiles recorded in other areas such as the Iberian Palaeomargin and other central Europe basins.
Keywords: Lofer cyclothems; bivalve assemblages; Middle Triassic; Lújar Unit; Granada province.
|
| |
RESUMEN
Este trabajo documenta el primer registro de vertebrados fósiles, concretamente de un reptil marino (sauropterygia), en las Zonas Internas de la Cordillera Bética. El desarrollo de plataformas epicontinentales en el Tetis durante la fragmentación de Pangea a partir del Triásico Inferior tardío favoreció la radiación de los sauropterigios, grupo de reptiles diápsidos marinos que incluye a los clados Placodontiformes, Pachypleurosauria, Nothosauroidea y Pistosauroidea. Los materiales triásicos del Complejo Alpujárride de las Zonas Internas de la Cordillera Bética, SE de España, incluyen los depósitos carbonatados correspondientes a las plataformas epicontinentales desarrolladas durante el Triásico medio-superior en el margen norte del Macizo Messomediterraneo. El intervalo estratigráfico seleccionado se encuentra en la Sierra de Lújar (Granada) y es una sucesión de 17,5 m de potencia con una alternancia de margas y calizas fosilíferas con abundante bioturbación y restos de macroinvertebrados. Se han identificado ciclos loferíticos con laminación a la base que cambia a un sedimento intensamente bioturbado con abundantes bivalvos hacia el techo del ciclo. El intervalo de estudio representa facies someras en un ambiente inter- o submareal como indican los ciclos loferíticos y la presencia de brechas carbonatadas en el techo de la secuencia, presumiblemente relacionadas con tormentas. Una costilla dorsal de 15 cm de longitud de un sauropterigio, probablemente un notosaurio, ha sido descubierta en esta serie en buen estado de conservación y sin evidencias de fragmentación, perforaciones o encostrantes. Este estudio muestra por primera vez la presencia de restos fósiles de reptiles marinos en el Complejo Alpujárride, reptiles que vivieron en aguas someras durante el Triásico al igual que se ha documentado en el Paleomargen Ibérico y en algunas cuencas triásicas de Europa central.
Palabras clave: Ciclos loferíticos; asociaciones de bivalvos; Triásico Medio; Unidad de Lújar; Provincia de Granada.
|
IntroductionTOP
During the fragmentation of Pangea, the extensional tectonic regime favoured the marine flooding of large areas and the development of epicontinental carbonate platforms in the Western Tethys (e.g. Martín-Rojas et al., 2009; Preto et al., 2010). The development of such epicontinental platforms, restricted to the Tethys Ocean, favoured the evolution and radiation of Sauropterygia, a diverse group of diapsid marine reptiles including Placodontiformes, Pachypleurosauria, Nothosauroidea, and Pistosauroidea since the late Lower Triassic (Rieppel, 2000; Motani, 2009).
The Triassic rocks of the Alpujarride Complex, in the Internal Zones of the Betic Cordillera, represent the carbonate deposition mainly developed during the Triassic in the northern margin of the Mesomediterranean Massif (Martín-Rojas et al., 2014; Ortí et al., 2017; Fig. 1). In the Alpujarride Complex, fossil assemblages are recognisable despite the intense metamorphism and dolomitization. These fossil assemblages are characterised by the record of macroinvertebrates such as bivalves, brachiopods, echinoderms, hermatypic corals, serpulids, as well as solenoporacean and dasycladacean algae (Braga, 1986; Martín & Braga, 1987a, b; Braga & Martín, 1987a, b), and less commonly nektonic organisms such as cephalopods (ammonoidea, coleoidea, and nautiloidea; Martín & Braga, 1987a, b). The interpreted palaeoenvironments are in general very shallow shelf deposits (Martín & Braga, 1987a; Somma et al., 2009). In such environments, at the Iberian Palaeomargin (Fig. 1), represented by the Muschelkalk facies, there are bivalves and brachiopod assemblages (Escudero-Mozo et al., 2015), but also marine reptiles remains (e.g. Niemeyer, 2002; Márquez-Aliaga et al., 2019).
|
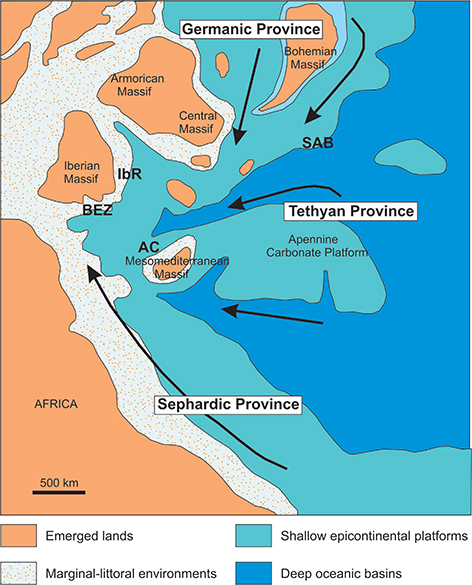 Figure 1.—Palaeogeographic map of the Ladinian times showing the advance of the influence of the different bioprovinces (modified after Márquez-Aliaga et al., 1986; Hirsch et al., 1987; Baeza-Carratalá et al., 2018) with indication of the areas represented today by different tectonic units. Note: AC, Alpujarride Complex (that includes La Forestal section); BEZ, Betic External Zones; IbR, Iberian Range; SAB, South Alpine Basin (that includes the Besano Fm). Figure 1.—Palaeogeographic map of the Ladinian times showing the advance of the influence of the different bioprovinces (modified after Márquez-Aliaga et al., 1986; Hirsch et al., 1987; Baeza-Carratalá et al., 2018) with indication of the areas represented today by different tectonic units. Note: AC, Alpujarride Complex (that includes La Forestal section); BEZ, Betic External Zones; IbR, Iberian Range; SAB, South Alpine Basin (that includes the Besano Fm).
|
|
Marine reptiles were diverse and abundant in Triassic marine environments of Europe, both in the Germanic Basin and in the Alpine Domain, forming part of the Germanic Bioprovince and the Tethyan Bioprovince (Fig. 1), summarised in Rieppel (2000), Motani (2009), and Diedrich (2012). At the Germanic Basin, from the Muschelkalk deposits, the most abundant reptiles in shallow marine sediments were Sauropterygia (e.g. Rieppel, 2000; Klein et al., 2015). In the Alpine Triassic, in addition to Sauropterygia, Ichthyosauria and Thalattosauria also occur (e.g. Müller, 2005; Dalla Vecchia, 2006, 2008; Kolb et al., 2011). The Besano Formation from the Monte San Giorgio (Southern Alps of Switzerland and Italy; Fig. 1) is especially rich in marine reptiles (e.g. Tschanz, 1989; Müller et al., 2005; Beardmore & Furrer, 2016; summarised in Rieppel, 2019).
The record of marine reptiles is relatively rich along the Iberian Palaeomargin, with Germanic facies (outcropping today in the Iberian Range and the Betic External Zones; Fig 1), but most of the reported remains consist of isolated elements (e.g. Sanz, 1976, 1983a, b; Alafont, 1992; Sanz et al., 1993; Niemeyer, 2002; Quesada & Aguera González, 2005; Fortuny et al., 2011; Reolid et al., 2014a; Miguel Chaves et al., 2015, 2016, 2017; Márquez-Aliaga et al., 2019). Considering the presence of marine reptile remains in shallow deposits from Germanic facies as well as the record of sauropterygians in the Alpine facies from other domains, it results interesting that there were not fossils reported from the Alpujarride Complex, the carbonate platform developed in the northern margin of the Mesomediterranean Massif (Fig. 1).
The objective of this work is to report the first record of sauropterygians from the Internal Zones of the Betic Cordillera, more exactly from the Middle Triassic of the Alpujarride Complex. In addition, this work aims to reconstruct the palaeoenvironment inhabited by these reptiles based on the analysis of lithofacies, microfacies, trace fossils, and macroinvertebrate assemblages.
Geological settingTOP
The study section, La Forestal (36º52’17”N, 3º26’56”W), is located in the Sierra de Lújar, Granada, SE Spain (Fig. 2). The Sierra de Lújar relief is part of the Alpujarride Complex, in the Betic Internal Zones (Fig. 2). The Alpujarride Complex consists of a Palaeozoic basement with micaschists and quartzites, and a cover made up of a lower Permian – Anisian formation with phyllite and quartzite, and an upper Middle Triassic to Upper Triassic carbonate formation, which includes a thin Jurassic deposit at the top (e.g. Delgado et al., 1981; Braga & Martin, 1987a; Sanz de Galdeano & López Garrido, 2014). Different tectonic units of the Alpujarride Complex are overthrusting after the Alpine Orogeny resulting in nappes. The Alpujarride Complex underwent different folding and metamorphic phases (Azañón & Goffé, 1997; Sanz de Galdeano & López Garrido, 2014). The metamorphic degree is also variable among different units.
The studied La Forestal outcrop is located in the Lújar Unit (Sanz de Galdeano & López-Garrido, 2014). According to Alonso-Chaves et al. (2004), six members constitute the Triassic carbonates of the Lújar Unit:
| • |
Member 1: Marly-limestones and dolostones (≈ 500 m, Lower Ladinian) |
| • |
Member 2: Fossiliferous marly-limestones (≈ 150 m, Lower to Middle Ladinian) |
| • |
Member 3: Cherty limestones (≈ 60 m, Middle Ladinian) |
| • |
Member 4: Limestones and dolostones (≈ 350 m, Upper Ladinian) Member 5: Marly-limestones (≈ 130 m, Carnian)Member 6: Dolostones (> 250 m, Norian) |
The studied interval represents shallow facies from intertidal to offshore facies of the Member 2 (Fossiliferous marly-limestone Member) of the Meta-carbonate Formation of the Lújar Unit.
MethodsTOP
The vertebrate remain was found by a hiker who contacted the university. The fieldwork involved the in-situ recognition of the main facies and sedimentary structures in a stratigraphic interval of 17.5 m thickness documented in a stratigraphic profile (Fig. 3). The documentation also included panorama photographs of the outcrop as well as close-ups of distinctive features. Finally, the different facies were sampled for microfacies analysis on thin sections. The microfacies analysis was portrayed on 12 thin-sections, stored in the Department of Geology of University of Jaén. The carbonates were classified according to Dunham (1962) and Embry & Klovan (1972). Seven marl samples were washed and sieved for studying the microfossil content but they were barren. These samples were analysed in the Laboratório Nacional de Energia e Geologie (Mamede de Infesta, Portugal) for palynomorphs with biostratigraphic purposes but no remains were found.
|
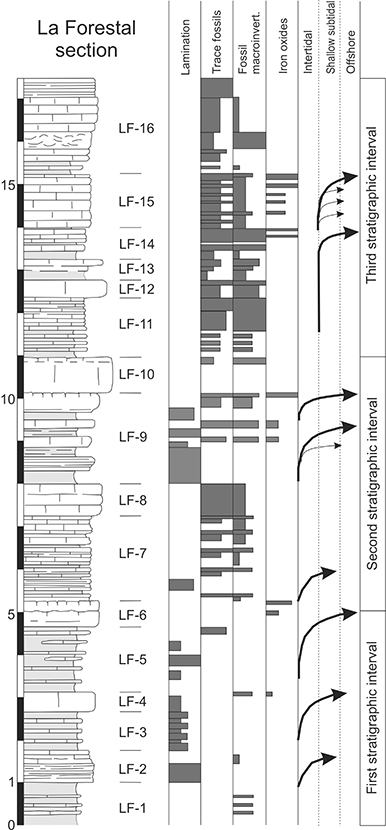 Figure 3.—Stratigraphic column of the La Forestal section with indication of sedimentary features observed in the field and thin sections as well as the differentiated stratigraphic intervals and identified deepening sequences (see text). Figure 3.—Stratigraphic column of the La Forestal section with indication of sedimentary features observed in the field and thin sections as well as the differentiated stratigraphic intervals and identified deepening sequences (see text).
|
|
The fossil reptile bone is now displayed in the Aula Museo de Paleontología of the University of Granada (Spain).
ResultsTOP
LithofaciesTOP
The La Forestal section includes part of the fossiliferous marly-limestone of the Member 2 of Alonso-Chaves et al. (2004). There are three parts differentiated according to the sedimentary structures and the presence of trace fossils and macroinvertebrates (Figs. 4 and 5). Limestones dominate over marly layers (Figs. 3 and 4) and a general trend toward increasing carbonate content is recognisable throughout the section (Fig. 3).
|
 Figure 4.—Field aspects of the La Forestal section. A. Lowermost part of the section with pink and yellow marls and laminated limestones. B. First and second stratigraphic intervals including yellow and pink marls and thin-bedded limestones. C. Field view of the upper part of the second stratigraphic interval that finishes at the top of the thick limestone bed LF-10. D. Thin-bedded limestones of the base of the third stratigraphic interval. The fossil bone was recovered from LF-13. E. Pink limestones of levels LF-14 rich in thin-shelled bivalves. F. Typical sedimentary sequence of La Forestal section with a laminated lower part and bioturbated upper part rich in fossil bivalves (example from LF-7). Figure 4.—Field aspects of the La Forestal section. A. Lowermost part of the section with pink and yellow marls and laminated limestones. B. First and second stratigraphic intervals including yellow and pink marls and thin-bedded limestones. C. Field view of the upper part of the second stratigraphic interval that finishes at the top of the thick limestone bed LF-10. D. Thin-bedded limestones of the base of the third stratigraphic interval. The fossil bone was recovered from LF-13. E. Pink limestones of levels LF-14 rich in thin-shelled bivalves. F. Typical sedimentary sequence of La Forestal section with a laminated lower part and bioturbated upper part rich in fossil bivalves (example from LF-7).
|
|
|
 Figure 5.—Fossil and ichnofossil assemblages. A. Top of the LF-7 with Pseudocorbula gregaria and secondarily Unionites and Homomya. B. Moulds of Pseudocorbula gregaria from the top of LF-7. C. Top of the limestone bed LF-9 rich in Umbostrea (arrows). D. Ferruginous top of the bed LF-10 rich in bivalve remains (mainly Limea, Costatoria, and Homomya) and an ammonite fragment (arrow). E. Dense bioturbation of Helminthopsis in most of the beds LF-15. F. Top of a bed from LF-16 thin-bedded limestones with abundance of bivalves (Limea, Homomya, Costatoria, Unionites, and Umbostrea) and small gastropods. Figure 5.—Fossil and ichnofossil assemblages. A. Top of the LF-7 with Pseudocorbula gregaria and secondarily Unionites and Homomya. B. Moulds of Pseudocorbula gregaria from the top of LF-7. C. Top of the limestone bed LF-9 rich in Umbostrea (arrows). D. Ferruginous top of the bed LF-10 rich in bivalve remains (mainly Limea, Costatoria, and Homomya) and an ammonite fragment (arrow). E. Dense bioturbation of Helminthopsis in most of the beds LF-15. F. Top of a bed from LF-16 thin-bedded limestones with abundance of bivalves (Limea, Homomya, Costatoria, Unionites, and Umbostrea) and small gastropods.
|
|
The lower five meters of the studied interval consists of an alternance of laminated marls and limestones (Figs. 3, 4A, B). The limestone intervals are thin bedded (5 to 15 cm thick) and made up of a mudstone with usually well-preserved lamination, mainly at the base of the beds (Fig. 4F, 6A, B) and trace fossils toward the top (Fig. 6C). Some layers are wackestones (locally packstones) including fragments of thin-shelled bivalves, gastropods and recrystallised benthic foraminifera such as in the bed LF-1 (Fig. 6D). The beds composing LF-2 display well-developed lamination is in (Fig. 3). Marly beds have a characteristic yellowish to pink colour (Fig. 4A, B). The top of some beds is rich in lumps and moulds of small bivalves (Fig. 5), and secondarily coal remains (< 2 cm). Trace fossils at the lower part of the studied section are scarce, only Rhizocorallium is locally present in limestone beds of LF-5. The upper part of this stratigraphic interval ends in a thick carbonate bed (LF-6) with some layers characterised by ferruginous tops with common bivalves and unbranched horizontal trace fossils with irregular meandering corresponding to Helminthopsis.
|
 Figure 6.—Microfacies from the La Forestal section. A and B. Laminated mudstones (beds LF-7 and LF-9). C. Mudstone with trace fossils and scarce bivalves and lumps (top of the bed LF-5). D. Wackestone of bioclasts, mainly bivalve fragments (bed LF-4). E. Packstone of recrystallised bivalves (bed LF-11). F. Alternance of laminated mudstones and packstones of bivalves in some cases showing erosive base (bed LF-11). Figure 6.—Microfacies from the La Forestal section. A and B. Laminated mudstones (beds LF-7 and LF-9). C. Mudstone with trace fossils and scarce bivalves and lumps (top of the bed LF-5). D. Wackestone of bioclasts, mainly bivalve fragments (bed LF-4). E. Packstone of recrystallised bivalves (bed LF-11). F. Alternance of laminated mudstones and packstones of bivalves in some cases showing erosive base (bed LF-11).
|
|
The second part of the section (5.7 m thick) has an increased carbonate content, abundant trace fossils, and benthic macroinvertebrates (Fig. 3, 4C). This interval consists of two thickening-upwards sequences of around 250 cm each, composed of thin-bedded limestones (5-10 cm thick) changing upward into thick limestone beds (70 cm and 80 cm thick in each sequence, LF-8 and LF-10 in Figure 2). The thin-bedded limestones at the base are laminated mudstone in contrast with the thick beds at top, which are densely bioturbated (Helminthopsis and secondarily Thalassinoides) wackestone rich in fossil bivalves (Fig. 5A, B). Among the thin-bedded limestones, thin marly layers occur. The thick limestone beds are highly bioturbated (Helminthopsis) and the top is rich in bivalves and iron oxides (Fig. 5C, D).
The third stratigraphic interval (6.6 m thick) contains non-laminated limestones with abundant trace fossils, bivalves, and common iron oxides at bed tops (Fig. 5E, F). Marly layers are very scarce. Most of the interval is a thin-bedded mudstone with intercalated bioclastic packstone layers with erosive bases (Fig. 6E and F). Bivalves are the main component of the bioclastic packstone. The top of the section consists of several fine-breccia layers with abundant intraclasts corresponding to fragmented (re-elaborated) trace fossils.
Fossil assemblagesTOP
Fossils in the study section are mainly bivalves belonging to Pseudocorbula gregaria, Costatoria sp., Limea sp., Homomya sp., Pseudoplacunopsis flabellum, Unionites sp. and Umbostrea sp., and small moulds of gastropods (Fig. 5). The species Pseudocorbula gregaria is dominant. A fragment of an indeterminate small ammonite was also recorded (bed LF-10). Small gastropods and echinoderms are recognisable in thin section. Costatoria and Unionites are preserved as recrystallized shells. Pseudocorbula gregaria appears at the top of some levels as dense accumulations mainly preserved as inner moulds (Fig. 5B).
A long curved fossil bone, corresponding to a dorsal rib of a sauropterygian, occur in the bed LF-13 at the lower part of the third stratigraphic interval. The bone occurred at the top of a horizon with Helminthopsis. It is well preserved without evidence of original fragmentation, borings, or encrustations. This rib is 14.98 cm long and the width varies from 9.7 to 24.1 mm (Fig. 7), with the thickest section in the distal region. The rib presents relative pachyostosis related to the compactness of the bone and the strong distal expansion. A thin neck (9.7 mm in diameter) with a low ridge in the dorsal side connects the rib head (proximal region). The head of the rib is swollen and shows a concave facet of articulation with vertebrae. The posterior third of the rib is slightly curved close to the position of the vertebrae. The internal surface is flat (ventrally flat, Fig. 7A) and the external surface of the rib is convex (dorsally convex, Fig. 7B), both striate to the distal region. The convexity increases to the proximal region in relation to the presence of a low ridge with a progressive development of triangular and finally pseudo-pentagonal cross-section (Fig. 7C).
|
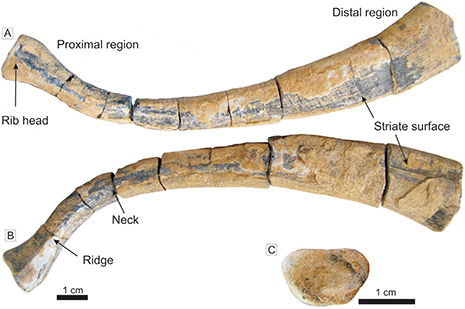 Figure 7.—Fossil rib of sauropterygian from the top of the bed LF-13. A. Internal view. B. External view. C. Articulation facet of the rib head. Figure 7.—Fossil rib of sauropterygian from the top of the bed LF-13. A. Internal view. B. External view. C. Articulation facet of the rib head.
|
|
DiscussionTOP
Palaeoenvironmental interpretationTOP
The laminated mudstones at the lower part of the section were deposited in a shallow environment, sometimes densely colonised by tracemakers doing subhorizontal burrows corresponding to Helminthopsis (less commonly to Thalassinoides). Helminthopsis was interpreted as grazing trails produced by deposit feeders, probably polychaetes (Bromley, 1996) that proliferated when sedimentation rate was reduced. The dense concentration and the small size of these trace fossils indicate large populations of tracemakers, and most likely an opportunistic behaviour. In addition, the dense concentration could depend on the time available for colonization the bottom. The recorded trace fossils commonly occur in yellowish horizons resembling omission surfaces. The local record of Rhizocorallium indicates reduced sediment input and omission-surface and firmground development (e.g., Cotillon et al., 2000; Worsley & Mørk, 2001; Rodríguez-Tovar et al., 2007; Reolid et al., 2014b). The presence of pebbly mudstones at bed tops is related to reworking that are associated to high-energy processes.
The base of the second interval (Fig. 3) is characterised by the record of laminated limestones with tops densely colonised by small subhorizontal trace fossils of Helminthopsis (and scarce Thalassinoides), indicating the opportunist colonization of the bottom when sedimentation rate decreased. The elementary sequences clearly display an upward increase in carbonate content and with densely burrowed tops that may represent gradual changes in the consistence of the substrate. The preservation of the burrows is an indicator of substrate consistence (Taylor et al., 2003), and consequently facilitates reconstruction of the sedimentation rate or depositional breaks. The record of horizons rich in trace fossils and iron oxides represents the transition from softground to firmground conditions. The tops of the elementary sequences with high concentrations of Pseudocorbula indicate a colonization of very shallow softground by infauna and the subsequent colonization of firmground (or hardground) by sessil epifauna.
The sedimentary features such as lamination are indicative of a shallow intertidal to subtidal environment whereas the densely bioturbated levels distinctive of subtidal to offshore facies. The sequences, of decimetric scale within beds and also of metric scale in thickening upward sequences, beginning with laminated limestones, followed by bioturbated limestones and occasionally finishing with fossiliferous horizons with iron oxides can be interpreted as deepening upward sequences (Fig. 3). The tops of these sequences are indicative omission surfaces or a hardgrounds, which include fragmentation and reworking of burrows resulting as lumps. These features resemble the flat-pebble breccias, typical product of storms in tidal carbonate areas. Assuming that subaerial early diagenetic processes were not responsible for lithification of the carbonate layers, the submarine early cementation driving to hardground formations was a necessary step in the origin of flat-pebbles included fragments of Helminthopsis. The reworking of these early cemented horizons was the source of flat-pebbles (Myrow et al., 2004). The flat-pebbles have been typically described from Palaeozoic, but also from Triassic (Wignall & Twitchett, 1999). The studied succession is constituted by typical sequences of Lofer cyclothems in the sense of Fischer (1964) that indicate deepening sequences from intertidal to subtidal in each sequence (Goldhammer et al., 1990; Satterley & Brandner, 1995). However, some type cycles of Lofer cyclothems have been reinterpreted as shoaling upward (Enos & Samankassou, 1998; Cozzi et al., 2005).
The bivalve association is similar to that of the Muschelkalk of the Iberian Ranges in the Landete Formation, Mediterranean Triassic domain, and at the base of the Cañete Formation, Levantine-Balearic Triassic domain (Escudero-Mozo et al., 2015). This association corresponds to the Sephardic Bioprovince that colonised the westernmost palaeogeographic regions of the Tethys (Márquez-Aliaga et al., 1986; Pérez-López et al., 1991), which currently constitute the Subbetic and Prebetic domains. This assemblage also colonised many areas of the Alpujarride Domain (López-Garrido et al., 1997). The association (Pseudocorbula, Costatoria, Limea, Homomya, Pseudoplacunopsis and Unionites) is characterised by low diversity and is often composed of dominant cosmopolitan specimens in the Alpine/Germanic facies (Palaeotethys) related to shallow and/or restricted environments. Senkowiczowa (1985) interpreted these cosmopolitan species as related to muddy bottoms, which is congruent with their occurrence in mudstones in La Forestal section. These genera of bivalves were pioneer taxa and probably lived under stressful environmental conditions (Escudero-Mozo et al., 2015). Unionites is indicative to marine and brackish waters (Márquez-Aliaga et al., 1986; Reolid & Reolid, 2017). The bivalve assemblage is consistent with the stressful conditions related to the tidal environment.
Sauropterygians in the Alpine Domain of SE SpainTOP
The proposed sedimentary environment is congruent with the habitat of marine reptiles such as basal sauropterygians, however during the Late Triassic some sauropterygians were adapted to pelagic environments. From the taphonomic point of view, the record of an isolated but well-preserved rib of a nothosaur is probably due to decoupling and transport of bones. The good preservation indicates short transport distances, para-autochthonous remain. In addition, there are not taphonomic features that evidence the activity of scavengers or long exposition on the sea-bottom before burial (such as potential record of borings or encrustations by sessile benthic organisms on the bone).
The features of the rib (the compactness of the bone, the distal expansion of the rib and the rib head connected by a thin neck with a low ridge in the dorsal side, slightly curved in its posterior third, among other features) indicate this corresponds to Eosauropterygia. Placodontia are excluded because they are not pachyostotic in the dorsal ribs (see more details of ribs of Placodontia in Diedrich, 2010; Klein & Scheyer, 2014; Klein et al., 2019). In the case of Paraplacodus, this is also excluded due to dorsal ribs of this genus present posteriorly directed uncinated processes (Klein & Sichelschmidt, 2014). In addition, placodontids do not have the characteristic distal expansion of the rib here described and a thin neck does not connect the rib head. However, the striated distal end resemble the dorsal rib of Pararcus diependroeki of the Muschelkalk of Netherland (Klein & Scheyer, 2014). Pachypleurosauria are also excluded because they do not present a rib morphology similar to the studied fossil remain and they are small-size sauropterygians (e.g. Rieppel, 2019).
Comparison of distal region of the rib with the same costal interval in typical Nothosauroidea such as Nothosaurus and Lariosaurus evidences that these genera maintain a more constant narrow diameter than the studied specimen does. The shape of the rib, with distal expansion, also resemble those of Paludidraco multidentatus a simosaurid recorded in the Upper Triassic of Central Spain (Miguel Chaves et al., 2018). The holotype exposed in the Museo de Paleontología de Castilla-La Mancha (Spain) shows a relatively constricted proximal region versus a strongly robust and expanded distal with relative degree of pachyostosis. In addition, the size proportions of the dorsal ribs of the Paludidraco holotype are within the size of the studied fossil rib.
The comparison with different genera of eosauropterygians (mainly Nothosauroidea such as Lariosaurus, Nothosaurus, “Paranothosaurus”, Simosaurus and Paludidraco, see Rieppel, 2000; Diedrich, 2012; Miguel Chaves et al., 2018) allow us for interpreting a total length of the specimen between 2.5 and 3.2 m, according to the length of the studied rib. The genus Lariosaurus is relatively common in some areas of the western shoreline of the Tethys with Alpine facies, for example in the Besano Formation (Southern Alps of Switzerland and Italy; see Rieppel, 2019). However, the recorded rib does not belong to Lariosaurus, a small-size Eosauropterygian with absence of the distal expansion. The estimated length are indicative of Nothosaurus, “Paranothosaurus”, and Simosaurus, but according to the morphology of the studied rib, the simosaurid Paludidraco seems to be a plausible candidate. However, only with an isolated rib, the reptile can exclusively be assigned to an indeterminate member large-size Eosauropterygian.
The occurrence of sauropterygians in the Member 2 of the Lújar Unit of the Alpujarride Complex (Fossiliferous marly-limestone Member) can be considered as the equivalent in the Alpine facies of the record of sauropterygians in Ladinian Muschelkalk facies of the External Zones of the Betic Cordillera, (e.g. Niemeyer, 2002). The record of marine reptiles in Triassic marine sediments of Europe is diverse and abundant in the Germanic Basin as well as in the Alpine Triassic (Rieppel, 2000; Motani, 2009; Diedrich, 2012). Along the Iberian palaeomargin, with Germanic facies, the record of marine reptiles is relatively rich but most of the reported remains consist of isolated elements, commonly reworked (see Sanz, 1976, 1983a, b; Alafont, 1992; Fortuny et al., 2011; Miguel Chaves et al., 2015, 2016, 2017; Márquez-Aliaga et al., 2019). The marine reptile fauna of northeastern Iberia includes diverse Sauropterygia with the nothosaur Lariosaurus balsami being the most common (Sanz et al., 1993; Rieppel & Hagdorn, 1998). In the south-eastern Iberian Palaeomargin, including the Tabular Cover and the External Zones of the Betic Cordillera, marine reptiles are represented by the nothosaurians Nothosaurus, Simosaurus, and Lariosaurus, by the placodonts Psephosaurus and Henodus, and by the Protorosauria Tanystropheus (Niemeyer, 2002, Reolid et al., 2014a). Those sauropterygian are the geographically nearer to the specimen studied in this work, but the south Iberian Palaeomargin and the Mesomediterranean Massif palaeomargin were separated by a marine corridor (Fig. 1). This fact gives relevance to this first record from a different palaeogeographic area, being the first record of a saurpterygian in the Alpine Domain of SE Spain.
ConclusionsTOP
This is the first report of fossil vertebrate remains of marine reptiles in the Triassic of the Alpujarride Complex, in spite of the extensive outcrops of carbonates. This is probably due to intense crushing, dolomitization, and locally metamorphism of the carbonates of the Alpujarride Complex. The studied sequence belongs to the Ladinian of the Lújar Unit.
The studied stratigraphic interval is 17.5 m thick of a succession of marls, laminated limestones and bioturbated limestones rich in fossil macroinvertebrates. Lofer cyclothems occur and display lamination at the base and intense bioturbation and abundant bivalves towards the top, locally with omission surfaces. There are deepening upward sequences that ranges from inter- to subtidal environments. Fossil bivalve fauna (mainly Pseudocorbula, and secondarily Costatoria, Limea, Homomya, Pseudoplacunopsis, Umbostrea, and Unionites) confirms this environmental interpretation.
The fossil bone corresponds to a well-preserved 15-cm long dorsal rib of an indeterminate member of large-size Eosauropterygian, probably a Nothosauroidea of the Family Simosauridae. This fossil appears in a similar environmental context than those reported from Iberian Palaeomargin in the Prebetic and in the Iberian Range.
ACKNOWLEDGEMENTSTOP
JR research was supported by the Juan de la Cierva Program Ministerio de Ciencia, Innovación y Universidades). MR research was supported by his own salary. The authors would like to thank the Buxus Association (Motril, Granada) for helping during the fieldwork and Prof. Ana Márquez-Aliaga for helping with the comparison of studied fossil bivalves with the faunas of the Iberian Palaeomargin. We also want to thank Prof. Juan Carlos Braga for his valuable comments on an early version of the manuscript. We would like to thank the reviewers Carlos De Miguel Chaves (UNED) and Dennis Voeten (Uppsala University) for their helpful comments.
ReferencesTOP
| ○ |
Alafont, L.S. (1992). Notosaurios y Placodontos (Reptilia) del Triásico Medio de Bienservida-Villarrodrigo. Instituto de Estudios
Albacetenses, Diputación de Albacete, Serie 1, 60, Albacete, Spain, 131 pp.
|
| ○ |
Alonso-Chaves, F.M.; Andreo, B.; Azañón, J.M.; Balanya, J.C.; Booth-Rea, G.; Crespo, A.; Delgado, F.; Estévez, A.; García-Casco,
A.; García-Dueñas, V.; López-Garrido, A.C.; Martín-Algarra, A.; Orozco, M.; Sánchez-Gómez, M.; Sánchez-Navas, A.; Sanz de
Galdeano, C. & Torres-Rondalla, R.I. (2004). Sucesiones litológicas, petrología y estratigrafía (del Complejo Alpujárride).
In: Geología de España (Vera, J.A., Ed.), SGE-IGME, Madrid, 409–411.
|
| ○ |
Azañón, J.M. & Goffé, B. (1997). Ferro- and magnesiocarpholite assemblages as record of high-P, low-T metamorphism in the
Central Alpujarrides, Betic Cordillera (SE Spain). European Journal of Mineralogy, 9: 1035–1051. https://doi.org/10.1127/ejm/9/5/1035 |
| ○ |
Baeza-Carratalá, J.F.; Pérez-Valera, F. & Pérez-Valera, J.A. (2018). The oldest post-Paleozoic (Ladinian, Triassic) brachiopods
from the Betic Range, SE Spain. Acta Palaeontologica Polonica, 63: 71–85. https://doi.org/10.4202/app.00415.2017 |
| ○ |
Beardmore, S.R. & Furrer, H. (2016). Preservation of Pachypleurosauridae (Reptilia; Sauropterygia) from the Middle Triassic
of Monte San Giorgio, Switzerland. Neues Jahrbuch fur Geologie und Palaontologie-Abhandlungen, 280: 221–240. https://doi.org/10.1127/njgpa/2016/0578 |
| ○ |
Braga, J.C. (1986). Upper Triassic Dasycladaceans in the Sierra de Baza (Alpujarride Complex, Betic Cordillera, SE Spain).
Neues Jahrbuch fur Geologie und Palaontologie Monatshefte, 1986/4: 239–247.
|
| ○ |
Braga, J.M. & Martín, J.M. (1987a). Sedimentación cíclica lagunar y bioconstrucciones asociadas en el Trías Superior alpujárride.
Cuadernos de Geología Ibérica, 11: 459–473.
|
| ○ |
Braga, J.C. & Martín, JM. (1987b). Distribución de las algas dasycladáceas en el Trías Alpujárride. Cuadernos de Geología
Ibérica, 11: 475–489.
|
| ○ |
Bromley, R.G. (1996). Trace fossils: Biology, taphonomy and applications. Chapman and Hall, London, 378 pp. |
| ○ |
Cotillon, P.; Banvillet, M.; Gaillard, C.; Grosheny, D. & Olivero, D. (2000). Les surfaces à Rhizocorallium de l’Aptien inférieur sur la bordure méridionale du bassin Vocontien (France Sud-Est), marqueurs de dynamiques locales; leur
relation avec un événement anoxique global. Bulletin de la Sociètè Géologique de France, 17: 229–238. https://doi.org/10.2113/171.2.229 |
| ○ |
Cozzi, A.; Hinnov, L.A. & Hardie, L.A. (2005). Orbitally forced Lofer cycles in the Dachstein Limestone of the Julian Alps
(northeastern Italy). Geology, 33: 789–792. https://doi.org/10.1130/G21578.1 |
| ○ |
Dalla Vecchia, F.M. (2006). A new sauropterygian reptile with plesiosaurian affinity from the Later Triassic of Italy. Rivista
Italiana di Paleontologia e Stratigrafia, 112: 207–225.
|
| ○ |
Dalla Vecchia, F.M. (2008). First record of Simosaurus (Sauropterygia, Nothosauroidea) from the Carnian (Late Triassic) of Italy. Rivista Italiana di Paleontologia e Stratigrafia,
114: 273–285.
|
| ○ |
Delgado, f.; Estévez Rubio, A.; Martín, J.M. & Martín Algarra, A. (1981). Observaciones sobre la estratigrafía de la Formación Carbonatada de los Mantos Alpujárrides (Cordillera Bética). Estudios geológicos, 37 (1–2): 45–58. |
| ○ |
Diedrich, C.G. (2010). Palaeoecology of Placodus gigas (Reptilia) and other placodontids - Middle Triassic macroalgae feeders in the Germanic Basin of central Europe - and evidence
for convergent evolution with Sirenia. Palaeoeography, Palaeoclimatology, Palaeocology 285: 287–306. https://doi.org/10.1016/j.palaeo.2009.11.021 |
| ○ |
Diedrich, C.G. (2012). The Middle Triassic marine reptile biodiversity in the Germanic Basin, in the centre of the Pangaean
world. Central European Journal of Geosciences, 4: 9–46. https://doi.org/10.2478/s13533-011-0060-0 |
| ○ |
Dunham, R.J. (1962). Classification of Carbonate Rocks According to Depositional Textures. In: Classification of carbonate rocks: a symposium (Ham, W.E. Ed.). American Association of Petroleum Geologists, pp. 108�121. |
| ○ |
Embry, A.F. & Klovan, J.E. (1972). Absolute water depth limits of Late Devonian paleoecological zones. Geologische Rundschau, 61(2): 672�686. |
| ○ |
Enos, P. & Samankassou, E. (1998). Lofer cyclothems revisited (late Triassic, northern Alps, Austria). Facies, 38: 207–227.
https://doi.org/10.1007/BF02537366 |
| ○ |
Escudero-Mozo, M.J.; Márquez-Aliaga, A.; Goy, A.; Martín-Chivelet, J.; López-Gómez, J.; Márquez, L.; Arche, A.; Plasencia,
P.; Pla, C.; Marzo, M. & Sánchez-Fernández, D. (2015). Middle Triassic carbonate platforms in Eastern Iberia: Evolution of
their fauna and palaeogeographic significance in the western Tethys. Palaeogeography, Palaeoeclimatology, Palaeoecology, 417:
236–260. https://doi.org/10.1016/j.palaeo.2014.10.041 |
| ○ |
Fischer, A.G. (1964). The Lofer cyclothems of the Alpine Triassic. Kansas Geological Survey Bulletin, 169: 107–149. |
| ○ |
Fortuny, J.; Bolet, A.; Sellés, A.G.; Cartanyà, J. & Galobart, A. (2011). New insights on the Permian and Triassic vertebrates
from the Iberian Peninsula with emphasis on the Pyrenean and Catalonian basins. Journal of Iberian Geology, 37: 65–86.
|
| ○ |
Goldhammer, R.K.; Dunn, P.A. & Hardie, L.A. (1990). Depositional cycles, composite sea-level changes, cycle stacking patterns,
and the hierarchy of stratigraphic forcing: examples from Alpine Triassic platform carbonates. Geological Society of America
Bulletin, 102: 535–562. https://doi.org/10.1130/0016-7606(1990)102<0535:DCCSLC>2.3.CO;2 |
| ○ |
Hirsch, F.; Márquez-Aliaga, A. & Santisteban, C. (1987). Distribución de moluscos y conodontos del tramo superior del Muschelkalk
en el Sector Occidental de la Provincia Sefardí. Cuadernos de Geología Ibérica, 11: 799–814.
|
| ○ |
Klein, N. & Scheyer, T.M. (2014). A new placodont sauropterygian from the Middle Triassic of the Netherlands. Acta Palaeontologica
Polonica, 59: 887–902. https://doi.org/10.4202/app.2012.0147 |
| ○ |
Klein, N. & Sichelschmidt, O.J. (2014) Remarkable dorsal ribs with distinct uncinated processes from the early Anisian of
the Germani Basin (Winterswijk, The Netherlands). Neues Jarbuch Paläontologische Abhandlungen, 271: 307–314. https://doi.org/10.1127/0077-7749/2014/0391 |
| ○ |
Klein, N.; Voeten, D.F.A.E.; Lankamp, J.; Bleeker, R.; Sichelschmidt, O.J.; Liebrand, M.; Nieweg, D.C. & Sander, P.M. (2015).
Postcranial material of Nothosaurus marchicus from the Lower Muschelkalk (Anisian) of Winterswijk, The Netherlands, with remarks on swimming styles and taphonomy. Paläontologische
Zeitzfrich, 89: 961–981. https://doi.org/10.1007/s12542-015-0273-5 |
| ○ |
Klein, N.; Canoville, A. & Houssaye, A. (2019). Microstructure of vertebrae, ribs, and gastralia of Triassic sauropterygians
- New insights into the microanatomical processes involved in aquatic adaptations of marine reptiles. The Anatomical Record,
302: 1770–1791. https://doi.org/10.1002/ar.24140 |
| ○ |
Kolb, C.; Sánchez-Villagra, M.R. & Scheyer, T.M. (2011). The palaeohistology of the basal ichthyosaur Mixosaurus Baur, 1887 (Ichthyopterygia, Mixosauridae) from the Middle Triassic: Palaeobiological implications. Comptes Rendus Palevol,
10: 403–411. https://doi.org/10.1016/j.crpv.2010.10.008 |
| ○ |
López-Garrido, A.C.; Pérez-López, A. & Sanz de Galdeano, C. (1997) Présence de faciès Muschelkalk dans des unités alpujarrides
de la région de Murcie (Cordillère Bétique, sud-est de l’Espagne) et implications paléogéographiques. Comptes Rendus Academie
Sciences Paris, 324: 647–654.
|
| ○ |
Márquez-Aliaga, A.; Hirsch, F. & López-Garrido, A.C. (1986). Middle Triassic bivalves from the Hornos-Siles Formation (Sephardic
Province, Spain). Neues Jahrbuch für Geologie und Paläonologie, Abhandlungen, 173: 201–227.
|
| ○ |
Márquez-Aliaga, A.; Klein, N.; Reolid, M.; Plasencia, P.; Villena, J.A. & Martínez-Pérez, C. (2019). An enigmatic marine reptile,
Hispaniasaurus cranioelongatus (gen. et sp. nov.) with nothosauroid affinities from the Ladinian of the Iberian Range (Spain). Historical Biology, 31: 223–233.
https://doi.org/10.1080/08912963.2017.1359264 |
| ○ |
Martín, J.M. & Braga, J.C. (1987a). Alpujarride carbonate deposits (southern Spain): marine sedimentation in a Triassic Atlantic.
Palaeogeography, Palaeoclimatology, Palaeoecology, 59: 243–260. https://doi.org/10.1016/0031-0182(87)90083-6 |
| ○ |
Martín, J.M. & Braga, J.C. (1987b). Bioconstrucciones del Anisiense-Ladiniense en el Trías Alpujárride. Cuadernos de Geología
Ibérica, 11: 421–444.
|
| ○ |
Martin-Rojas, I.; Somma, R.; Delgado, F.; Estévez, A.; Iannace, A.; Perrone, V. & Zamparelli, V. (2009). Triassic continental rifting of Pangaea: direct evidence from the Alpujarride carbonates, Betic Cordillera, SE Spain. Journal of the Geological Society, 166(3), 447–458. https://doi.org/10.1144/0016-76492008-091 |
| ○ |
Martín-Rojas, I.; Somma, R.; Estévez, A.; Delgado F. & Zamparelli, V. (2014). La plataforma triásica alpujárride (Zonas Internas
de la Cordillera Bética, España). Revista de la Sociedad Geológica de España, 27: 63–78.
|
| ○ |
Miguel Chaves, C. De; Pérez-García, A.; Cobos, R.; Royo-Torres, F.; Ortega, F. & Alcalá L. (2015). A diverse Late Triassic
tetrapod fauna from Manzanera (Teruel, Spain). Geobios, 48: 479–490. https://doi.org/10.1016/j.geobios.2015.09.002 |
| ○ |
Miguel Chaves, C. De; García-Gil, S.; Ortega, F.; Sanz, J.L. & Pérez-García, A. (2016). First Triassic tetrapod (Sauropterygia,
Nothosauridae) from Castilla y León: evidence of an unknown taxon for the Spanish record. Journal of Iberian Geology, 42:
29–38. https://doi.org/10.5209/rev_JIGE.2016.v42.n1.51210 |
| ○ |
Miguel Chaves, C. De; Ortega, F. & Pérez-García, A. (2017). The eosauropterygian fossils from the Middle Triassic of Canales
de Molina (Guadalajara, Spain). Journal of Iberian Geology, 43: 129–138. https://doi.org/10.1007/s41513-017-0011-9 |
| ○ |
Miguel Chaves, C. De; Ortega, F. & Pérez-García, A. (2018). New highly pachyostotic nothosauroid interpreted as a filter-feeding
Triassic marine reptile. Biology Letters, 14: 20180130. https://doi.org/10.1098/rsbl.2018.0130 |
| ○ |
Motani, R. (2009). The evolution of marine reptiles. Evolution: Education and Outreach, 2: 224–235. https://doi.org/10.1007/s12052-009-0139-y |
| ○ |
Müller, J. (2005). The anatomy of Askeptosaurus italicus from the Middle Triassic of Monte San Giorgio, and the interrelationships of thalattosaurs (Reptilia, Diapsida). Canadian
Journal of Earth Sciences, 42: 1347–1367. https://doi.org/10.1139/e05-030 |
| ○ |
Myrow, P.M.; Tice, L.; Archuleta, B.; Clark, B.; Taylors, J.F. & Ripperdan, R.L. (2004). Flat-pebble conglomerate: its multiple
origins and relationship to metre-scale depositional cycles. Sedimentology, 51: 973–996. https://doi.org/10.1111/j.1365-3091.2004.00657.x |
| ○ |
Niemeyer, J. (2002). Invertebraten und Vertebraten aus dem Muschelkalk von Siles (Jaén), Spanien. Münstersche Forschungen
zur Geologie und Paläontogie, 94: 1–99.
|
| ○ |
Ortí, F.; Pérez-López, A. & Salvany, J.M. (2017). Triassic evaporites of Iberia; sedimentological and palaeogeographical implications
for the western Neotethys evolution during the Middle Triassic-Earliest Jurassic. Palaeogeography, Palaeoclimatology, Palaeoecology,
471: 157–180. https://doi.org/10.1016/j.palaeo.2017.01.025 |
| ○ |
Pérez-López, A. (1991). El Trías de facies germánicas del sector central de la Cordillera Bética. PhD Thesis, Universidad
de Granada, 400 pp.
|
| ○ |
Preto, N.; Kustatscher, E. & Wignall, P.B. (2010). Triassic climates: State of the art and perspectives. Palaeogeography,
Palaeoclimatology, Palaeoecology, 290: 1–10. https://doi.org/10.1016/j.palaeo.2010.03.015 |
| ○ |
Quesada, J.M. & Aguera González S. (2005). Descripción del primer ejemplar de Ceresiosaurus (Reptilia: Sauropterygia), hallado en la Península Ibérica en el Ladiniense (Triásico Medio) de Mont-Ral-Alcover (Tarragona).
Estudios Geológicos, 61: 247–269. https://doi.org/10.3989/egeol.05613-667 |
| ○ |
Reolid, M.; Marok, A. & Lasgaa, I. (2014b). Taphonomy and ichnology: tools for interpreting a maximum flooding interval in
the Berriasian of Tlemcen Domain (Western Tellian Atlas, Algeria). Facies, 60: 905–920. https://doi.org/10.1007/s10347-014-0413-5 |
| ○ |
Reolid, M.; Pérez-Valera, F.; Benton, M.J. & Reolid, J. (2014a). Marine flooding event in continental Triassic facies identified
by a nothosaur and placodont bone bed (South Iberian Paleomargin). Facies, 60: 277–293. https://doi.org/10.1007/s10347-013-0360-6 |
| ○ |
Reolid, J. & Reolid, M. (2017). Traces of floating archosaurs: an interpretation of the enigmatic trace fossils from the Triassic
of the Tabular Cover of Southern Spain. Ichnos, 24: 222–233. https://doi.org/10.1080/10420940.2016.1265524 |
| ○ |
Rieppel, O. (2000). Sauropterygia I. In: Encyclopedia of Paleoherpetology (Wellnhofer, P., Ed.), Volume 12A, Dr. Friedrich
Pfeil Verlag, München, 134 pp.
|
| ○ |
Rieppel, O. (2002). Feeding mechanics in Triassic stem group sauropterygians: the anatomy of a successful invasion of Mesozoic
seas. Zoological Journal Linnean Society 135: 33–63. https://doi.org/10.1046/j.1096-3642.2002.00019.x |
| ○ |
Rieppel, O. & Hagdorn, H. (1998). Fossil reptiles from the Spanish Muschelkalk (Montral and Alcover, Province Tarragona).
Historical Biology, 13: 77–97. https://doi.org/10.1080/08912969809386575 |
| ○ |
Rieppel, O. (2019). Mesozoic sea dragons: Triassic marine life from the Ancient tropical lagoon of Monte San Giorgio. Indiana
University Press, 256 pp. https://doi.org/10.2307/j.ctvd58t86 |
| ○ |
Rodríguez-Tovar, F.J.; Pérez-Valera, F. & Pérez-López, A. (2007). Ichnological analysis in high-resolution sequence stratigraphy:
The Glossifungites ichnofacies in Triassic successions from the Betic Cordillera (southern Spain). Sedimentary Geology, 198: 293–307. https://doi.org/10.1016/j.sedgeo.2007.01.016 |
| ○ |
Sanz, J.L. (1976). Lariosaurus balsami (Sauropterygia, Reptilia) de Estaeda (Huesca). Estudios Geológicos, 32: 547–567.
|
| ○ |
Sanz, J.L. (1983a). Los Nothosaurios (Reptilia, Sauropterygia) Españoles. Estudios Geológicos, 39: 193–215. |
| ○ |
Sanz, J.L. (1983b). Consideraciones sobre el género Pistosaurus. El suborden Pistosauria (Reptilia, Sauropterygia). Estudios Geológicos, 39: 451–458.
|
| ○ |
Sanz, J.L.; Alafont, L.S. & Moratalla, J.J. (1993). Triassic reptile faunas from Spain. In: Evolution, Ecology and Biogeography
of the Triassic Reptiles (Mazin, J.M. & Pinna, G., Eds.), Paleontol Lombarda NS, 2: 153–164.
|
| ○ |
Sanz de Galdeano, C. & López-Garrido, A.C. (2014). Structure of the Sierra de Lujar (Alpujarride Complex, Betic Cordillera).
Estudios Geológicos, 70: e005. https://doi.org/10.3989/egeol.41491.290 |
| ○ |
Satterley, A.K. & Brandner, R. (1995). The genesis of Lofer Cycles of the Dachstein Limestone, Northern calcareous Alps, Austria.
Geologische Rundschau, 84: 287–292. https://doi.org/10.1007/BF00260441 |
| ○ |
Somma, R.; Martín-Rojas, I.; Delgado, F.; Estévez, A.; Iannace, A.; Perrone, V. & Zamparelli, V. (2009). Facies and facies
association distribution on the Triassic carbonate platform of the Alpujarride Complex (Betic Internal Zone, SE Spain). Geogaceta,
47: 81–84.
|
| ○ |
Taylor, A.; Goldring, R. & Gowland, S. (2003). Analysis and application of ichnofabrics. Earth-Science Reviews, 60: 227–259.
https://doi.org/10.1016/S0012-8252(02)00105-8 |
| ○ |
Tschanz, K. (1989). Lariosaurus buzzii n. sp. from the Middle Triassic of Monte San Giorgio (Switzerland) with comments on the classification of nothosaurs. Palaeontographica.
Abteilung A, Paläozoologie, Stratigraphie, 208(4-6): 153–179.
|
| ○ |
Wignall, P.B. & Twitchett, R.J. (1999). Unusual intraclastic limestones in Lower Triassic carbonates and their bearing on
the aftermath of the end-Permian mass extinction. Sedimentology, 46: 303–316. https://doi.org/10.1046/j.1365-3091.1999.00214.x |
| ○ |
Worsley, D. & Mørk, A. (2001). The environmental significance of the trace fossil Rhizocorallium jenense in the Lower Triassic
of western Spitsbergen. Polar Research, 20: 37–48. https://doi.org/10.1111/j.1751-8369.2001.tb00037.x |