Nuevos datos sobre las faunas marinas del Eoceno medio-superior de Navarra (área surpirenaica occidental). Revisión de los fósiles de la colección Ruiz de Gaona.
New data on the marine faunas from the middle–upper Eocene of Navarre (Western South Pyrenean Area). The fossils of the Ruiz de Gaona collection revisited.
Humberto Astibia1,*, José-Carmelo Corral2, Germán Álvarez-Pérez3, Miguel Ángel López Horgue1, Aitor Payros1
1Departamento de Geología, Facultad de Ciencia y Tecnología, Universidad del País Vasco/Euskal Herriko Unibertsitatea, Apartado de correos 644, 48080 Bilbao. ORCID IDs: https://orcid.org/0000-0001-5690-0030; https://orcid.org/0000-0002-9703-0948; https://orcid.org/0000-0001-9549-4119
2Museo de Ciencias Naturales de Álava/Arabako Natur Zientzien Museoa, Siervas de Jesús, 01001 Vitoria-Gasteiz. ORCID ID: https://orcid.org/0000-0001-9493-3693
3Departament d’Estratigrafia, Paleontologia i Geociències Marines. Universitat de Barcelona. Martí i Franqués s/n. 08028 Barcelona. ORCID ID: https://orcid.org/0000-0003-0053-4881
*Corresponding author: humberto.astibia@ehu.eus
| |
RESUMEN
Durante el Eoceno el área pirenaica pudo desempeñar un papel importante como encrucijada paleobiogeográfica entre los dominios del Tetis occidental y el Atlántico norte. Sin embargo, sus faunas marinas están todavía poco documentadas. En este trabajo se presentan los resultados de un primer estudio sobre los macrofósiles marinos del Eoceno medio–superior (Luteciense–Bartoniense–Priaboniense) de Navarra de la colección paleontológica de Máximo Ruiz de Gaona, investigador cuyos estudios sobre la geología y paleontología de este territorio constituyen una referencia histórica obligada. Los fósiles descritos provienen de las formaciones margosas del Eoceno de la cuenca de Pamplona y de la Formación Calizas de Urbasa-Andia y niveles suprayacentes, en el área de Estella-Urbasa, al oeste de la falla de Pamplona. Algunos de los especímenes estudiados provienen de afloramientos ya desaparecidos, circunstancia que aumenta el interés de la conservación y estudio de esta colección. Además, los datos paleontológicos del área de Estella–Urbasa son casi del todo novedosos. Este estudio ha permitido describir 24 taxones de macrofauna marina. Siete de ellos ─Cyclolitopsis patera, Funginellastraea barcelonensis, Pycnodonte rarilamella, Metacrinus sp., Triplacidia sp., además de un briozoo Cyclostomatida y un anélido Serpulidae indeterminados─ son nuevos para el registro fósil de Navarra. Las asociaciones macropaleontológicas descritas aquí y en otros trabajos precedentes son poco diversas si se las compara con las coetáneas de otras regiones geológicas europeas. Además de posibles condicionamientos ecológicos, se plantea la existencia de sesgos de muestreo y, sobre todo de sesgos tafonómicos, en concreto, la pérdida de las especies de invertebrados con esqueletos aragoníticos, como principal explicación de este hecho.
Palabras clave: faunas marinas; Eoceno; Navarra; colección Ruiz de Gaona.
|
| |
ABSTRACT
During Eocene times the Pyrenean area may have played an important role as a paleobiogeographic link between the Western Tethys and the North Atlantic domains. However, its marine faunas are still poorly documented. This paper presents the results of a first study on the mid–upper Eocene (Lutetian to Priabonian) marine macrofossils of Navarre included in the paleontological collection of Máximo Ruiz de Gaona, a researcher whose studies on the geology and paleontology of this territory constitute an obligatory historical reference. The described fossils come from the Eocene marly formations of the Pamplona basin and the Urbasa-Andia limestone Formation and overlying deposits, in the Estella-Urbasa area, west of the Pamplona fault. Some of the specimens studied come from outcrops that do not exist any longer. This fact increases the interest of conservation and the study of this collection. The paleontological data from the Estella-Urbasa area are almost entirely new. This study has allowed the description of 24 taxa of marine macrofauna. Seven of them ─Cyclolitopsis patera, Funginellastraea barcelonensis, Pycnodonte rarilamella, Metacrinus sp., Triplacidia sp., in addition to an indeterminate Cyclostomatida bryozoan and an indeterminate Serpulidae annelid─ are new to the fossil record of Navarre. The macropaleontological associations described in this and previous works are not as diverse as those from other coeval European geological regions. In addition to possible ecological conditions, sampling biases and, above all, taphonomic biases, specifically the loss of invertebrate species with aragonite skeletons, arise as the main explanation.
Keywords: marine fauna; Eocene; Navarre; Ruiz de Gaona collection.
|
Introducción y objetivosTOP
AntecedentesTOP
Durante el Eoceno medio el área pirenaica pudo desempeñar un papel importante como encrucijada paleobiogeográfica entre los altamente biodiversos dominios marinos del Tetis occidental y el Atlántico norte y, asimismo, haber albergado un número importante de especies endémicas (Astibia et al., 2018). Sin embargo, las faunas marinas del Eoceno medio pirenaico todavía están poco documentadas. En el área norpirenaica, obras clásicas como las de d’Archiac (1846, 1850a), Rouault (1850), Bouillé (1873, 1876), Boussac (1911), Cossmann (1921) y O’Gorman & Cossmann (1923), sobre los moluscos y otros invertebrados del “nummulítico” de la costa vasca y de Aquitania, apenas han sido actualizadas.
En lo que respecta al área surpirenaica, un número limitado de estudios, tales como los de Pisera & Busquets (2002), sobre esponjas, Álvarez-Pérez (2009), sobre corales, Villalta Comella (1956), De Renzi (1971, 1996), Abad (2001) y Dominici & Kowalke (2007, 2014), sobre moluscos, y Brzobohatý & Nolf (2011) y Díaz-Berenguer et al. (2018), sobre vertebrados, se han centrado en las faunas del Eoceno de Cataluña y Aragón.
En la región surpirenaica occidental, algunos de los datos paleontológicos más antiguos provienen del geólogo francés Carez (1854–1932), quien describió los extensos depósitos margosos de la cuenca de Pamplona (Carez, 1881) y mencionó en las cercanías de la capital navarra la abundancia de un pequeño bivalvo que llamó Plicatula pamplonensis [Dimya pamplonensis (Carez 1881), Calzada & Astibia 1996]. Marquina (1908) estudió el Eoceno en Navarra, presentó una demarcación de zonas con nummulítidos y citó la presencia de tallos fósiles de crinoideos (“Pentacrinus”) y conchas de Dimya pamplonensis en las margas de la cuenca de Pamplona. Posteriormente, Ruiz de Gaona (1947a), Mendizábal & Ruiz de Gaona (1949), y Ruiz de Gaona & Colom (1950) describieron el contenido micropaleontológico (foraminíferos) de estas margas, sugirieron su edad Bartoniense y mencionaron la presencia en algunos lugares de ricas asociaciones macrofaunísticas. Años más tarde, la microfauna de estas series margosas fue estudiada por Puigdefábregas (1975) y Alameda et al. (1993). Una serie de publicaciones más recientes se han centrado en el bivalvo Dimya pamplonensis (Calzada & Astibia 1996), el anélido Rotularia spirulaea (Elorza & Astibia, 2017, 2018) y las asociaciones fósiles de tortugas y sirenios (Astibia et al. 1999, 2005, 2006; Buffrénil et al., 2008), esponjas y corales (Astibia et al., 2014), braquiópodos (Bitner et al. 2016), moluscos gasterópodos y bivalvos (Astibia et al., 2018) e icnofósiles (Payros et al., 2000; Astibia et al., 2007, 2017). La mayoría de estas publicaciones no constituyen todavía sino trabajos de carácter general, realizadas con el objetivo de sentar unas bases para estudios futuros más detallados que, de llevarse a cabo, permitan un conocimiento más preciso de la biodiversidad marina durante el Eoceno en el golfo pirenaico.
Máximo Ruiz de GaonaTOP
Puede afirmarse que Máximo Ruiz de Gaona (Espronceda, 1902 – Pamplona-Iruñea, 1971) fue el fundador de la Paleontología en Navarra y uno de los principales paleontólogos que emergió en el estado español en los difíciles tiempos de la última postguerra. Máximo Ruiz de Gaona ingresó en la Orden de las Escuelas Pías en 1918 y se ordenó sacerdote hacia 1924. Con gran mérito personal, se licenció en Ciencias Naturales con más de 50 años. Su obra científica se materializó en más de una treintena de publicaciones, un número notable, dados el escaso tiempo que tuvo durante años para dedicarse a la investigación y los limitados medios materiales de los que dispuso para ello. Los trabajos de Ruiz de Gaona constituyen hoy una referencia histórica obligada para quienes estudian la geología y paleontología del Cretácico y Paleógeno pirenaico. Su labor investigadora se centró en la micropaleontología de foraminíferos, pero su inquieta mente y, en particular, su interés por las ciencias del pasado, le llevaron a otros campos, como la geología, la paleontología de vertebrados y la prehistoria (Astibia et al., 1996; Latasa, 1996).
Favorecido por sus estancias en varios centros escolapios (Barbastro, Estella, Tamarite de Litera, Bera, Molina de Aragón, Jaca, Tolosa y Pamplona; véase Latasa, 1996), Ruiz de Gaona dejó a su fallecimiento en el Colegio de los Escolapios de Pamplona-Iruñea una importante colección de micro y macrofósiles, sobre todo del Cretácico y Paleógeno surpirenaicos, incluida la región vasco-cantábrica (también hay algunas muestras recogidas y clasificadas por él en el Colegio de los Escolapios de Bilbao). Hoy en día, la Colección Ruiz de Gaona (CRG) se guarda en los Fondos de Arqueología del Departamento de Cultura, Deporte y Juventud (Sección de Arqueología, Servicio de Patrimonio Histórico) del Gobierno de Navarra. Aunque las condiciones de almacenaje y conservación de la colección son buenas, la misma necesitaría un riguroso trabajo de documentación y catalogación. La CRG incluye numerosos materiales no publicados y, como es lógico por el paso de los años, aquellos que sí lo fueron, precisan ser revisados. Muchos fósiles provienen de lugares hoy desaparecidos y/o nuevos desde un punto de vista paleontológico, hechos que aumentan el interés del inventariado y estudio de esta colección (véase p. e. Salazar et al., 2017).
ObjetivosTOP
El objetivo general de este trabajo es realizar una primera descripción de los macrofósiles, principalmente de invertebrados, del Eoceno medio y superior (Luteciense, Bartoniense y Priaboniense) de Navarra de la CRG, en sus contextos geológico, tafonómico y sistemático. Objetivos concretos son: (1) revisar y describir sucintamente diversos fósiles del Luteciense y Bartoniense de la cuenca de Pamplona, algunos correspondientes a especies mencionadas pero no descritas en sus trabajos por el ilustre escolapio y otros de especies no conocidas en trabajos precedentes sobre esta zona de estudio y (2) estudiar un material fósil del Eoceno de la sierra de Urbasa y de la localidad de Anderatz, en Tierra Estella-Estellerria. En este segundo caso, se trata en la mayor parte de datos paleontológicos inéditos.
El presente trabajo supone una revisión de una pequeña parte de la CRG, sobre la que, en relación con sus fósiles de invertebrados, sólo existen los precedentes de los artículos de López-Horgue & Bodego (2017), en el que se describe uno de los taxones aquí incluidos, el crustáceo Harpactoxanthopsis, y de Elorza & Astibia (2018), sobre el anélido Rotularia spirulaea, de las localidades navarras de Anderatz y la sierra de Urbasa. En cuanto a los fósiles de vertebrados de la CRG, se han publicado hasta el momento los trabajos de Astibia et al. (1987), sobre los mamíferos del yacimiento del Mioceno de las Yeseras de Monteagudo, y Arlegi et al. (2018), sobre los mamíferos del Pleistoceno de la cueva de Koskobilo, ambas localidades también ubicadas en Navarra. Con este nuevo artículo se pretende contribuir a la valoración científica y cultural de la CRG, y recordar, una vez más, la figura de su entusiasta creador, cerca de tres cuartos de siglo después de sus trabajos más notorios sobre el Eoceno de Navarra. Desde un punto de vista paleontológico, este texto pretende ser una pequeña aportación al conocimiento del registro fósil del Eoceno de Navarra, en el contexto general del conocimiento de las faunas marinas del Eoceno pirenaico y de la evolución de la biodiversidad marina global durante esa época geológica.
Encuadre geológicoTOP
Los fósiles estudiados provienen de la Montaña y Zona Media de la Navarra peninsular, en el área surpirenaica occidental; en concreto de Tierra Estella-Estellerria y la sierra de Urbasa y de la cuenca de Pamplona-Iruñerria (Fig. 1a). En el Eoceno, el área surpirenaica se estructuró como una cuenca de antepaís orientada al noroeste, adyacente a un orógeno en formación situado al norte. Durante la mayor parte del Eoceno la cuenca de antepaís constituyó una estrecha franja de mar abierta al Golfo de Bizkaia a unos 35°N de paleolatitud. Con el tiempo, los aportes sedimentarios provenientes de la erosión de los Pirineos emergentes fueron rellenando la cuenca. Durante el Eoceno medio y superior (Bartoniense y Priaboniense) grandes sistemas sedimentarios fluvio-deltaicos se extendieron por la cuenca de antepaís (Plaziat, 1981; Pujalte et al., 2002; Barnolas et al., 2004). Esta evolución está bien registrada en la zona de estudio.
|
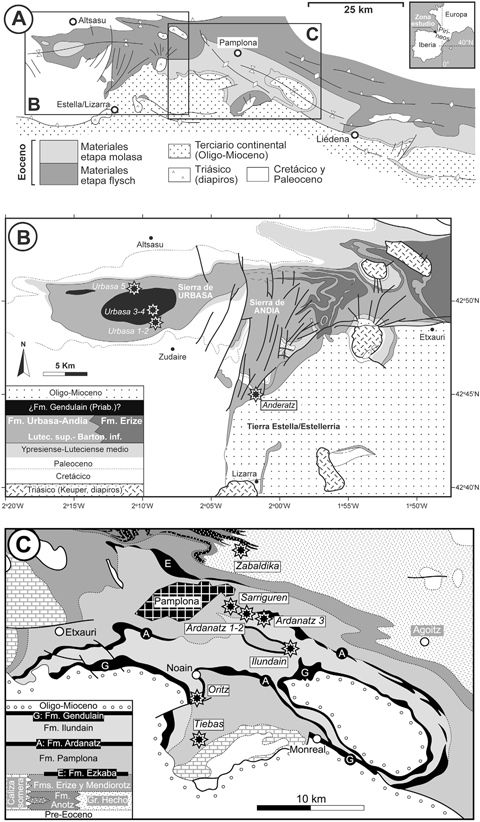 Fig. 1.— Mapas geológicos simplificados del Eoceno de la zona surpirenaica occidental (A), de la Cuenca de Pamplona y áreas circundantes (B), y de Tierra Estella y las sierras de Urbasa y Andia (C). La localización de los yacimientos citados en este trabajo está señalada con estrellas y sus nombres aparecen en cursiva. Fig. 1.— Mapas geológicos simplificados del Eoceno de la zona surpirenaica occidental (A), de la Cuenca de Pamplona y áreas circundantes (B), y de Tierra Estella y las sierras de Urbasa y Andia (C). La localización de los yacimientos citados en este trabajo está señalada con estrellas y sus nombres aparecen en cursiva.
|
|
Tierra EstellaTOP
Los fósiles de Tierra Estella provienen de la Formación Calizas de Urbasa-Andia (Payros, 1997; Payros et al., 2010) en el borde norte de la depresión de Estella-Lizarra (Fig. 1b). En esta formación predominan calcarenitas de rampa carbonatada marina somera de tipo foralgal dominada por tormentas, desarrollada bajo condiciones transgresivas, pero también incluye sedimentos margosos (Payros et al., 2010). Las Calizas de Urbasa-Andia abarcan desde el Luteciense superior hasta el Bartoniense, con niveles pertenecientes a las biozonas SBZ15, SBZ16 y SBZ17 de la biozonación de Serra-Kiel et al. (1998).
Sierra de UrbasaTOP
Algunos de los fósiles de la sierra de Urbasa también se incluyen en la Formación Calizas de Urbasa-Andia (unidades litológicas 263 y 268 del Mapa Geológico de Navarra 1:25.000, http://geologia.navarra.es/). Otros proceden de los niveles margosos con yeso suprayacentes a esta formación en la zona (unidad litológica 267 del Mapa Geológico de Navarra 1:25.000) (Fig. 1b). Según el Mapa Geológico de Navarra, la unidad 267 consta de margas bartonienses equivalentes a las Margas de Pamplona. Sin embargo, según criterios litológicos y sedimentológicos Payros (1997) y Payros et al. (2000) apuntaron su posible correlación con depósitos más jóvenes, como la Formación Gendulain del Eoceno superior (Priaboniense), definida en la cuenca de Pamplona.
Cuenca de PamplonaTOP
Los fósiles de la cuenca de Pamplona aquí descritos provienen casi en su totalidad de las formaciones Margas de Pamplona, Areniscas de Ardanatz y Margas de Ilundain, en el flanco norte del sinclinal de Aranguren-Itzaga (Fig. 1c). Sin embargo, un ejemplar procede de la localidad de Zabaldika, emplazada sobre la Formación de Margas y Calcarenitas de Anotz en su transición a la Formación Erro (Grupo Hecho). La Formación Anotz se depositó en un talud submarino carbonatado, a unos 500 m de profundidad (Payros et al., 2007), entre las biozonas P9 (Ypresiense superior) y P11 (Luteciense medio) de Berggren et al. (1995) (zonas E7-E9 de Wade et al., 2011). La Formación Margas de Pamplona también se formó en aguas relativamente profundas (Ruiz de Gaona y Colom, 1950; Puigdefabregas, 1975; Astibia et al., 2005). La parte baja de la Formación Margas de Pamplona podría corresponder a la zona P14 de Berggren et al. (1995) (zona E13 de Wade et al., 2011) del Bartoniense, según la edad de las unidades infra y suprayacentes (Astibia et al., 2005). La Formación Areniscas de Ardanatz (también referidas en la literatura regional como areniscas de Zizur, Gazolaz, Tajonar, Tabar o Gongolaz) está formada por hiperpicnitas arenosas acumuladas en lóbulos de frente deltaico (Astibia et al. 1999, 2005, 2006, 2014). La Formación Areniscas de Ardanatz se interdigita lateral y verticalmente con la Formación Margas de Ilundain, interpretada como un conjunto de depósitos marinos restringidos y poco profundos (Astibia et al., 2014). En algunos puntos de la cuenca de Pamplona las Margas de Pamplona e Ilundain se suceden sin la intercalación de las areniscas de Ardanatz. Según Astibia et al. (1999), los fósiles de las Areniscas de Ardanatz se incluyen en la parte alta de la zona SBZ17 de Serra-Kiel et al. (1998), indicativa asimismo de una edad Bartoniense, ya supuesta a partir de su contenido micropaleontológico por Ruiz de Gaona (1947a) y Mendizábal & Ruiz de Gaona (1949).
Material y métodosTOP
Localidades fosilíferasTOP
Los fósiles estudiados provienen de los siguientes lugares:
| 1. |
Zabaldika (Esteribar). Junto al puente viejo de la carretera N-135 sobre el río Arga. Arcillas con niveles arenosos, parte superior del Miembro Calciclástico 3 de la Formación Anotz (Zona P11, Luteciense medio, de Berggren et al., 1995, zona E9 de Wade et al., 2011), en su transición lateral a la Formación Erro del Grupo Hecho (Payros et al., 2007).
|
| 2. |
Tiebas (Tebas-Muru Artederreta). Afloramiento difícil de localizar o seguramente desaparecido, debido a la construcción de infraestructuras viarias, urbanización y roturación del término. Cuenca de Pamplona, parte baja de la Formación Margas de Pamplona, Bartoniense.
|
| 3. |
Oritz (Valle de Elortz-Elortzibar). probablemente junto o en el término de Santa Cruz. Cuenca de Pamplona, parte baja de la Formación Margas de Ilundain, Bartoniense.
|
| 4. |
Sarriguren (Valle de Egüés-Eguesibar). La etiqueta no precisa más. Afloramiento desaparecido, ya que la práctica totalidad del término de esta localidad, hoy sede del ayuntamiento del valle, está roturado y urbanizado. Cuenca de Pamplona, parte alta (?) de la Formación Margas de Pamplona, Bartoniense.
|
| 5. |
Ardanatz 1 (Eguesibar). Afloramiento muy cercano al anterior, desaparecido por roturación del terreno, situado aproximadamente en el kilómetro 20 de la carretera local NA-2310 (desde Pamplona, pasando por la población de Sarriguren, a unos 2 kilómetros antes del desvío —NA-2315— que va a la pequeña localidad de Ardanatz). En las etiquetas de la CRG aparecen como “Ardanaz, km 2 antes del pueblo”. Cuenca de Pamplona, parte alta de la Formación Margas de Pamplona o parte baja de la Formación Margas de Ilundain, Bartoniense. Indicamos su pertenencia al Valle de Egüés-Eguesibar para diferenciar esta localidad de su homónima del cercano valle de Itzagaondoa.
|
| 6. |
Ardanatz 2 (Eguesibar). Afloramiento desaparecido, situado aproximadamente en el kilómetro 20,5 de la carretera local NA-2310 (desde Pamplona, pasando por la población de Sarriguren, a unos 2,5 kilómetros antes del desvío —NA-2315— que va a esta localidad). En las etiquetas de la CRG aparecen como “Ardanaz, km 2,5 antes del pueblo”. Cuenca de Pamplona, parte alta de la Formación Margas de Pamplona o parte baja de la Formación Margas de Ilundain, Bartoniense.
|
| 7. |
Ardanatz 3 (Eguesibar). Varios afloramientos de las cercanías de la población de este nombre, pertenecientes muy probablemente a las secciones AD1, AD2 y AD3 de Astibia et al. (2014). En las etiquetas de la colección RDG aparecen como “Ardanaz”, a veces con alguna indicación de localización poco precisa. Cuenca de Pamplona, transición entre las formaciones Areniscas de Ardanatz y Margas de Ilundain, Bartoniense.
|
| 8. |
Ilundain (Valle de Aranguren). Desconocemos la localización exacta de los afloramientos de donde provienen los materiales con esta indicación. Cuenca de Pamplona, parte alta de la Formación Margas de Pamplona o parte baja de la Formación Margas de Ilundain, Bartoniense.
|
| 9. |
Anderatz (Abartzuza). Las etiquetas sólo indican “Andéraz”, sin detallar más. Calcarenitas de la Formación Calizas de Urbasa-Andia. Esta formación abarca desde el Luteciense superior (SBZ15, biozonación de Serra-Kiel et al., 1998) hasta el Bartoniense (SBZ17), aunque los afloramientos de la zona de Abartzuza corresponden exclusivamente a SBZ17 (Payros et al., 2010).
|
| 10-14. |
Urbasa 1–5. Fósiles provenientes de afloramientos ubicados en la sierra de Urbasa, a lo largo de la carretera provincial Olatzagutia-Estella (NA-718), a la altura del kilómetro 23, margas y margocalizas de la parte superior (SBZ16-17?) de la Formación Urbasa-Andia (Payros et al., 2010), unidad cartográfica 268 del Mapa Geológico de Navarra 1:25.000, Bartoniense (Urbasa 1); kilómetro 24, margas y margocalizas, zona de límite entre las unidades cartográficas 268 (Formación Urbasa-Andia) y 267 del Mapa Geológico de Navarra 1:25.000, Bartoniense–Priaboniense (?) (Urbasa 2); kilómetros 24–25, margas, unidad cartográfica 267 del Mapa Geológico de Navarra 1:25.000, Priaboniense (?) (Urbasa 3); kilómetro 25, margas, unidad cartográfica 267 del Mapa Geológico de Navarra 1:25.000, Priaboniense (?) (Urbasa 4); y kilómetro 28, calizas margosas con discociclínidos y montículos locales de briozoos y esponjas, parte alta (SBZ17?) de la Formación Calizas de Urbasa-Andia (secuencia DS5 de Payros et al., 2010), unidad cartográfica 263 del Mapa Geológico de Navarra 1:25.000, Bartoniense (Urbasa 5). |
Hace más de medio siglo que Ruiz de Gaona (1947b) realizó un estudio del Eoceno de la sierra de Urbasa, recogiendo fósiles de foraminíferos (nummulítidos, alveolínidos, sorítidos y discociclínidos) en afloramientos situados a la altura de los kilómetros 21 y 30 de la carretera Olatzagutia-Estella. Desconocemos si fue entonces cuando también recogió los fósiles de invertebrados (esponjas, moluscos bivalvos, equinodermos equinoideos, artrópodos crustáceos y otros) que se encuentran en su colección y se incluyen en este estudio.
Hoy día, tanto en Anderatz, debido a las roturaciones agrícolas y el crecimiento de la vegetación en algunas zonas colindantes a los campos, como en los puntos de la sierra de Urbasa donde recolectó las muestras Ruiz de Gaona, seguramente hoy más cubiertas por la vegetación que entonces, apenas quedan superficies aflorantes prospectables. No obstante, la presencia de fósiles de macroforaminíferos, colonias de briozoos, Rotularia spirulaea, Pycnodonte brongniarti, fragmentos de bivalvos pectínidos, radiolas de equínidos, artejos de crinoideos y dientes de selacios, entre otros, en la localidad de Anderatz, y de fósiles de macroforaminíferos, briozoos y bivalvos (ostreidos y Spondylus sp.) en Urbasa, ha podido ser constatada a partir de observaciones recientes realizadas sobre el terreno por uno de los autores (H. A.) de este trabajo.
Preparación de las muestrasTOP
Los fósiles se han limpiado manualmente e introducido en bolsas individuales de polietileno, conservándose las cajas de cartón y etiquetas originales. Algunos especímenes se han preparado químicamente con hidróxido de potasio para eliminar el sedimento margoso adherido a los mismos. Se ha procedido asimismo al fotografiado de varios ejemplares, en algunos casos tras recubrimiento previo con un sublimado de cloruro amónico. Algunos de los ejemplares de menor tamaño se han fotografiado mediante microscopio electrónico de barrido (MEB). Las medidas de los fósiles se han tomado con calibre digital.
Abreviaturas y referencias bibliográficasTOP
CRG, colección Ruiz de Gaona; MGS, Museu Geològic del Seminari de Barcelona; MNHN, Muséum nátional d’Historire naturelle, París; NHM, Natural History Museum, Londres; H, altura (en los moluscos bivalvos diámetro umbo-paleal); A, anchura; D, diámetro; E, grosor; L, longitud (en los moluscos bivalvos diámetro antero-posterior); máx, máxima; mín, mínima; x, media; n, número de ejemplares medidos. Todas las medidas están en milímetros. Las cifras entre paréntesis indican dimensiones menores que las reales o aproximadas, debido al estado incompleto o fragmentario de muchos de los fósiles. En el listado de referencias bibliográficas no se incluyen las correspondientes a los autores de las categorías taxonómicas supragenéricas.
Paleontología sistemáticaTOP
Filo Porifera Grant, 1836
Clase Hexactinellida Schmidt, 1870
Guettardiscyphia thiolati(d’Archiac, 1846) o Pleuroguettardia iberica Pisera y Busquets, 2002
(Fig. 2a-b)
|
 Fig. 2.— Fósiles del Eoceno [Bartoniense–Priaboniense (?)] de Navarra de la Colección Ruiz de Gaona. Guettardiscyphia thiolati(d’Archiac, 1846) o Pleuroguettardia iberica Pisera y Busquets, 2002, CRG.11, vistas lateral (a) y oblicua (b), Ardanatz 3. Heteroscleromorpha indet., CRG.12, vistas superior (c), inferior (d) y lateral-oblicua (e), Ardanatz 1; CRG.59, vista lateral (f), Ardanatz 1. Placosmiliopsis bilobatus (d’Achiardi, 1868), CGR.13, vistas superior (g) y lateral (h), Ardanatz 3; CRG.14, vista lateral (i); CRG.61, sección en vista superior (j), Ardanatz 3. Cyclolitopsis patera (d’Achiardi, 1867), CRG.15, vistas lateral (k) e inferior (l), Ardanatz 3. Funginellastraea barcelonensis (Oppenheim, 1911), CRG.16, vistas lateral (m) e inferior (n), Ardanatz 3. Escalas a-b, g-n, 10 mm; c-f, 5 mm. Fig. 2.— Fósiles del Eoceno [Bartoniense–Priaboniense (?)] de Navarra de la Colección Ruiz de Gaona. Guettardiscyphia thiolati(d’Archiac, 1846) o Pleuroguettardia iberica Pisera y Busquets, 2002, CRG.11, vistas lateral (a) y oblicua (b), Ardanatz 3. Heteroscleromorpha indet., CRG.12, vistas superior (c), inferior (d) y lateral-oblicua (e), Ardanatz 1; CRG.59, vista lateral (f), Ardanatz 1. Placosmiliopsis bilobatus (d’Achiardi, 1868), CGR.13, vistas superior (g) y lateral (h), Ardanatz 3; CRG.14, vista lateral (i); CRG.61, sección en vista superior (j), Ardanatz 3. Cyclolitopsis patera (d’Achiardi, 1867), CRG.15, vistas lateral (k) e inferior (l), Ardanatz 3. Funginellastraea barcelonensis (Oppenheim, 1911), CRG.16, vistas lateral (m) e inferior (n), Ardanatz 3. Escalas a-b, g-n, 10 mm; c-f, 5 mm.
|
|
Material: Un fragmento proveniente de Ardanatz 3 (Eguesibar, Navarra), Formación Areniscas de Ardanatz, transición de la Formación Areniscas de Ardanatz a la Formación Margas de Ilundain (sección y nivel desconocidos, muy probablemente recogido en la parte baja de la sección AD3 de Astibia et al., 2014), Bartoniense.
Dimensiones: H = 34,1; A = 32,4
Descripción: El fósil corresponde a una esponja de forma alargada con tres pliegues o alas de la pared que le dan una sección transversal estrellada. Varios ósculos parietales se disponen a lo largo de los extremos o crestas de los pliegues. No hay ósculos en las partes laterales de las alas. Toda la superficie está cubierta por finas aberturas o poros, en algunas zonas siguiendo un patrón cuadrangular relativamente regular, pero en otras, por el contrario, la distribución de los poros es más irregular.
Observaciones: Astibia et al. (2014) han descrito fósiles similares de la misma localidad de Ardanatz (Eguesibar) que asignan a los géneros (y probables especies)
Guettardiscyphia [especie más probable Guettardiscyphia thiolati (d’Archiac, 1846)] y/o Pleuroguettardia (especie más probable Pleuroguettardia iberica Pisera & Busquets, 2002). Ambas especies son externamente muy similares, pero pertenecen a familias diferentes, Cribrospongiidae y Craticulariidae, respectivamente, que poseen diferentes patrones de canalización, irregular en la primera, cuadrangular o craticulárido, en la segunda. Con todo, en ambos taxones los patrones de canalización pueden cambiar o quedar alterados en algunas zonas del esqueleto y este hecho dificulta su diferenciación en el caso de disponer de material insuficiente (cf. Pisera & Busquets, 2002). Astibia et al. (2014) indican la posibilidad de que en la asociación de Ardanatz estén presentes ambas especies, pero la diferenciación se complica sin un estudio detallado, difícil de llevar a cabo por la pobre preservación y el estado fragmentario de la muestra disponible.
El material tipo de Guettardiscyphia thiolati —se desconoce su paradero— proviene del Eoceno de Rocher de Goulet, enclave cercano a la ciudad de Biarritz, en la costa vasca (área norpirenaica, cuenca de Aquitania). En la descripción original de la especie “Guettardia thiolati”, d’Archiac (1846: 197, pl. V, fig. 15, pl. VIII, figs. 5, 6, 7) menciona la disposición irregular de los poros o aberturas de los canales (“les cellules [=aberturas] ne paraissent pas d’ailleurs affecter une disposition symétrique régulière”), hecho que sugiere la pertenencia de dicho material al género Guettardiscyphia. Las ilustraciones del trabajo de d’Archiac (1846) muestran solamente la forma estrellada de los especímenes, pero no el detalle de sus paredes. Pleuroguettardia iberica ha sido definida en el Eoceno (Bartoniense–Priaboniense, Cascella & Dinarès-Turell, 2009) de la Formación Margas de Vic (cuenca del Ebro, Cataluña) (Pisera & Busquets, 2002: 335–337, figs. 7, 8 y 12). El género Pleuroguettardia también se menciona en el Eoceno Medio (Luteciense inferior) de Val di Chiampo, donde Matteucci & Russo (2005) asignan algunos especímenes a Pleuroguettardia aff. iberica y Pleuroguettardia sp. Guettardiscyphia thiolati está presente, asimismo, en las dos áreas anteriores.
Clase Demospongia Sollas, 1875
Heteroscleromorpha indet.
(Fig. 2c-f)
Material: Nueve ejemplares provenientes de Ardanatz 1 (Eguesibar, Navarra), parte alta de la Formación Margas de Pamplona o parte baja de la Formación Margas de Ilundain, Bartoniense.
Dimensiones: H = 4,7–12,2; A = 7,0–4,2
Descripción: Se trata de una esponja de pequeño tamaño, de forma globular o hemisférica achatada. Su superficie es finamente porosa y estriada, con canales o surcos profundos bifurcados dispuestos más o menos radialmente. En algunos ejemplares la base es circular y plana, con una marcada granulación que presenta una disposición reticulada bastante regular.
Observaciones: Esta esponja ya ha sido descrita por Astibia et al. (2014, Fig. 9a-f) como Lithistida sp. indet. A. Sus fósiles son comunes en la parte baja de la sección AD3 de Ardanatz (nivel AD3.1) y en la cercana sección de Badoztain (Margas de Ilundain, nivel BA1.2). También aparece en los afloramientos de la sección de Arraitza, a unos 25 km al oeste de Ardanatz, ubicada estratigráficamente unos 100 m por encima de la base de la Formación Margas de Ilundain (niveles ARR1.1 y ARR1.2) (Astibia et al., 2014). D’Archiac (1850a: 413, pl. 9) describe y figura una pequeña esponja que considera una nueva especie y denomina Tragos mamillatus, proveniente de la colección de M. Pratt (listada en d’Archiac, 1847) del Eoceno “nummulítico” de Biarritz, en la costa vasca del área norpirenaica (cuenca de Aquitania). Desconocemos el paradero de este material, aunque por su talla y morfología Heteroscleromorpha indet. podría asignarse a la misma especie. Sin embargo, la ausencia en los dos casos de un estudio detallado de sus elementos esqueléticos impide por el momento realizar determinaciones precisas.
Filo Coelenterata Frey y Leuckart, 1847
Clase Anthozoa Ehrenberg, 1834
Subclase Hexacorallia Haeckel, 1896
Orden Scleractinia Bourne, 1900
Familia Montlivaltiidae Dietrich, 1926
Género Placosmiliopsis Russo, 1979
Especie tipo: Placosmilia bilobata d’Achiardi, 1868
Placosmiliopsis bilobatus (d’Achiardi, 1868)
(Fig. 2g-j)
1868 Placosmilia bilobata – d’Achiardi: 25
1875 Placosmilia eocenica – d’Achiardi: 9
1901 Placosmilia eocaenica Reuss – Oppenheim: 72-73
1979 Placosmiliopsis bilobatus (d’Achiardi) − Russo: 61–62, pl. 8, fig. 1–2
Material: Seis ejemplares (uno de ellos probable forma anómala) provenientes de Ardanatz 3 (Eguesibar, Navarra), transición de la Formación Areniscas de Ardanatz a la Formación Margas de Ilundain, Bartoniense.
Dimensiones: H = 39,5–20,1; A = 51,1–29,2; E = 30,0–17,2
Descripción: Coral solitario de forma flabelada. Fijación por un pedúnculo basal. Cáliz elíptico. Fósula profunda, alargada en la dirección del eje mayor del cáliz. Los septos se disponen en seis ciclos, con simetría radial hexameral. S1 y S2 son gruesos y ropaloides. S3 son más finos y ligeramente más cortos. Los otros septos son más finos y cortos. Los márgenes distales de los septos son lisos. Las caras laterales de los septos son espinosas. La columnela es laminar, sinuosa y profunda. La pared es paraseptotecal en la superficie y septotecal en profundidad. Toda la superficie del esqueleto (corálum) está recorrida por una alternancia de costillas finas y gruesas que se corresponden con los septos. A medida que se desciende por la pared externa del pólipo, las costillas de los ciclos sexto, quinto y cuarto, desaparecen progresivamente. El resto recorren el pedúnculo basal de fijación al suelo. Endoteca vesicular bien desarrollada, sobre todo en la zona periférica. Esta especie presenta diversas anomalías: el cáliz puede ser circular, elíptico, en forma de ocho o del todo irregular; en sentido vertical, el pólipo puede ser corto o alargado y recto o curvado (ver Álvarez-Pérez & Busquets, 2012).
Observaciones: La presencia de fósiles de corales en el Bartoniense de Ardanatz ya fue indicada por Mendizábal & Ruiz de Gaona (1949) y Ruiz de Gaona & Colom (1950), sin que precisaran más en cuanto a su clasificación taxonómica. Los aquí descritos aparecen como Pattalophylia sp. y Petrophylliela sp. en las etiquetas originales de la CRG. En todos los casos la morfología de estos ejemplares es coincidente con la de varios fósiles asignados por Astibia et al. (2014) a la especie Placosmiliopsis bilobatus (d’Achiardi, 1868), provenientes de la parte alta de la sección AD3 de Ardanatz (Eguesibar) y de otros puntos no precisados de la misma localidad por los mismos autores. De distribución eocénica (“Ilerdiense”–Priaboniense), P. bilobatus está también presente en otras asociaciones de corales de la cuenca de antepaís surpirenaica (Álvarez-Pérez, 2009).
Familia Cyclolitidae d’Orbigny, 1851
Género Cyclolitopsis Reuss, 1874
Especie tipo: Cyclolites patera d’Achiardi, 1867
Cyclolitopsis patera (d’Achiardi, 1867)
(Fig. 2k-l)
1867 Cyclolites patera – d’Achiardi: 8.
1867 Cyclolites zignoi – d’Achiardi: 8.
1874 Cyclolitopsis patera – Reuss: 16, pl. 41, fig., 2–3.
1872 Cyclolites garnieri – Tournouër: 499, pl. 6, fig. 5.
1973 Funginellastraea garnieri – Barta-Calmus: 508, pl. 47, fig. 15–17.
Material: Dos ejemplares provenientes de Ardanatz 3 (Eguesibar, Navarra), transición de la Formación Areniscas de Ardanatz a la Formación Margas de Ilundain, Bartoniense.
Dimensiones: H = 12,2–(3,4); A = 28,3–11,0
Descripción: De los dos ejemplares, el de mayor tamaño (CRG.15) es un coral solitario de forma claramente patelada, cónica deprimida, con un corto pedúnculo basal de fijación al substrato. El cáliz es circular o elíptico, con la parte superior plana. Los septos se corresponden con las costillas (costo-septos) y están frecuentemente anastomosados. El borde superior es dentado. Los septos de los primeros órdenes son compactos mientras que los de los últimos órdenes son perforados. El grosor de los septos depende del orden al que pertenecen, siendo más gruesos los de los tres primeros ciclos, los cuales permanecen libres. Los septos del sexto ciclo se unen lateralmente a los del quinto ciclo y estos a los del cuarto ciclo. El centro del cáliz presenta una fosa calicinal profunda, carente de columnilla central. Los anillos concéntricos que conforman la epiteca no sobrepasan el número de doce. No posee ningún foraminífero (Discocyclinidae) en el centro. El segundo ejemplar es de menor talla y su forma es más bien discoidal. El área de fijación está truncada y no se ven restos ni de pedúnculo ni de ningún foraminífero. Las costillas son claramente visibles, al no estar recubiertas por la epiteca.
Observaciones: Los fósiles descritos se guardan con el nombre de Cycloseris andianensis, según la etiqueta original de Ruiz de Gaona. Definida en el Eoceno (Luteciense) del Véneto alpino (d’Achiardi, 1867), C. patera está presente en el Eoceno surpirenaico del área de Igualada (Cataluña) (Serra-Kiel et al., 2003). Su presencia era hasta ahora desconocida en el registro fósil del Eoceno de Navarra.
Familia Fungiidae Dana, 1846
Género FunginellastraeaAlloiteau, 1952
Especie tipo: Cycloseris patera Felix, 1909
Funginellastraea barcelonensis (Oppenheim, 1911)
(Fig. 2m-n)
1909 Cycloseris patera – Felix: 121, lám. 12, fig. 9–11.
1911 Cycloseris barcelonensis – Oppenheim: 336, fig. 5–6.
1952 Funginellastraea barcelonensis – Alloiteau: 345.
Material: Tres ejemplares provenientes de Ardanatz 3 (Eguesibar, Navarra). En la etiqueta original se indica “Ardanaz, SW de la cima”. Transición de la Formación Areniscas de Ardanatz a la Formación Margas de Ilundain, Bartoniense.
Dimensiones: H = 3,80–1,60; A = 12,3–7,4
Descripción: Coral solitario de forma discoidal, fijado a una discociclina que aparece en el centro de la cara basal. Posee una epiteca muy visible, formada por marcados anillos concéntricos.
Observaciones:Estas formas discoidales se han clasificado desde antiguo como pertenecientes a los géneros Cyclolites y Cycloseris. En la etiqueta original de la CRG los fósiles aquí descritos se asignan al segundo de los géneros. En Cataluña y Aragón se han descrito las especies Cyclolites borsonis Michelin, 1841, Cyclolites alpina d’Orbigny, 1850, Cyclolites andianensis d’Archiac, 1850, Cyclolites patera d’Achiardi 1867, Cyclolites garnieri Tournouër, 1872, Cyclolites heberti (Tournouër, 1872), Cyclolites rhomboideus Oppenheim, 1901, Cycloseris perezi Haime en d’Archiac 1850b y Cycloseris duplicicosta Oppenheim en Faura 1926. Felix (1909) describe una nueva especie en el Eoceno de la zona de Igualada, Cycloseris patera, aceptada inicialmente por Oppenheim (1910). Sin embargo, más tarde Oppenheim (1911) revisa los ejemplares recolectados y cree encontrar suficientes diferencias como para crear un nuevo género, que denomina Funginellastraea, y una nueva especie, que denomina Funginellastraea barcelonensis. Esta especie se encuentra en las cuencas surpirenaicas de Aragón y de Cataluña (Álvarez-Pérez, 1993, 2009) y hasta ahora no había sido citada en el Eoceno de Navarra.
Filo Brachiopoda Duméril, 1806
Clase Rhynchonellata Williams, Carlson, Brunton, Holmer & Popov, 1996
Orden Terebratulida Waagen, 1883
Superfamilia Cancellothyridoidea Thomson, 1926
Familia Cancellothyrididae Thomson, 1926
Subfamilia Cancellothyridinae Thomson, 1926
Género Terebratulina d’Orbigny, 1847
Especie tipo: Anomia retusa Linnaeus, 1758.
Terebratulina tenuistriata (Leymerie, 1846)
(Fig. 3a-d)
|
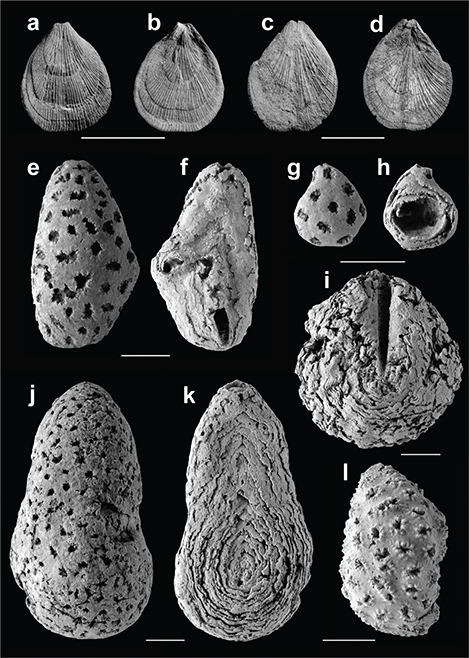 Fig. 3.— Fósiles del Eoceno [Bartoniense–Priaboniense (?)] de Navarra de la Colección Ruiz de Gaona. Terebratulina tenuistriata (Leymerie, 1846), CRG.17, vistas ventral (a) y dorsal (b), Ardanatz 1; CRG.18, vistas ventral (c) y dorsal (d), Ilundain. Cyclostomatida indet., CRG19, vistas anterior (e) y posterior (f), Urbasa 3; CRG.20, vistas anterior (g) y posterior (h), Urbasa 3; CRG.21, vista posterior (i), Urbasa 3; CRG.22, vistas anterior (j) y posterior (k), Urbasa 3; CRG.23 (sigla original: 1021), vista anterior (l), Urbasa 3. Escala 10 mm. Fig. 3.— Fósiles del Eoceno [Bartoniense–Priaboniense (?)] de Navarra de la Colección Ruiz de Gaona. Terebratulina tenuistriata (Leymerie, 1846), CRG.17, vistas ventral (a) y dorsal (b), Ardanatz 1; CRG.18, vistas ventral (c) y dorsal (d), Ilundain. Cyclostomatida indet., CRG19, vistas anterior (e) y posterior (f), Urbasa 3; CRG.20, vistas anterior (g) y posterior (h), Urbasa 3; CRG.21, vista posterior (i), Urbasa 3; CRG.22, vistas anterior (j) y posterior (k), Urbasa 3; CRG.23 (sigla original: 1021), vista anterior (l), Urbasa 3. Escala 10 mm.
|
|
1846 Terebratula tenuistriata – Leymerie: 363, pl. 15, fig. 11.
Sinonimias completas en Bitner & Müller (2017: 214–216).
Material:Tres ejemplares completos y tres en estado fragmentario provenientes de Ilundain (Aranguren, Navarra), parte alta de la Formación Margas de Pamplona o parte baja de la Formación Margas de Ilundain, Bartoniense. Diecinueve ejemplares de Ardanatz 1 (Eguesibar, Navarra), algunos aplastados y fracturados, parte alta de la Formación Margas de Pamplona o parte baja de la Formación Margas de Ilundain, Bartoniense.
Dimensiones: Ilundain: L = 16,2–11,0; A =14–8,4 / Ardanatz: L = 13,4–5,7; A = 11,0–4,4
Descripción: Braquiópodo micromórfico con contorno de la concha muy variable, desde sub-circular y sub-pentagonal hasta ovalado alargado. La superficie de la concha está cubierta por numerosas costillas finas que aumentan por intercalación y bifurcación. El foramen es redondeado, mesotírido a permesotírido con placas deltidiales pequeñas, triangulares y disjuntas.
Observaciones: La presencia de Terebratulina tenuistriata en el Eoceno de Navarra ya fue indicada por Ruiz de Gaona (1947a) y Mendizábal & Ruiz de Gaona (1949) en el Bartoniense de Ardanatz, “Tejería” (probablemente en el término de Mendillorri) y los Altos de Badoztain. Ruiz de Gaona & Colom (1950) añadieron a estas localidades la del término de Ezkoriz (indicado como “Escori”). En un reciente trabajo sobre los braquiópodos del Eoceno de la cuenca de Pamplona, Bitner et al. (2016), describen ejemplares de T. tenuistriata provenientes de las localidades de Badoztain (Formación Margas de Pamplona [?] – parte baja de la Formación Margas de Ilundain), Ardanatz y Aranguren (transición entre las formaciones Areniscas de Ardanatz y Margas de Ilundain) y Arraitza (Formación Margas de Ilundain). Se trata de la especie más común en el Eoceno de la zona y la única presente en la colección CRG.
T. tenuistriata es una especie común en el Eoceno de Europa, incluida la región pirenaica (Bitner, 2000), así como en el sur del Tetis (Bitner & Boukhary, 2012). También se ha reconocido en el Oligoceno inferior de Alemania (Bitner & Müller, 2015). En la CRG, entre los ejemplares de Ilundain los hay de mayor talla que los provenientes de Ardanatz. Al igual que los estudiados por Bitner et al. (2016), los de la CRG son en general menores que los del Eoceno inferior (Ilerdiense medio) de la sección de Campo (Huesca) (véase Bitner, 2000). Los ejemplares del afloramiento de Ilundain y algunos otros de Ardanatz son de mayor talla que los del Eoceno de Hungría (véase Bitner & Dulai, 2008).
Filo Bryozoa Ehrenberg, 1831
Clase Stenolaemata Borg, 1926
Orden Cyclostomatida Busk, 1852
Cyclostomatida indet.
(Fig. 3e-l)
Material: Diecisiete ejemplares provenientes de Urbasa 3 (sierra de Urbasa, Navarra), unidad cartográfica 267, Mapa Geológico de Navarra 1:25:000, Priaboniense (?).
Dimensiones: L = 54,0–9,5; A = 32,0–5,5; E = 22,4–3,6
Descripción:Zoario grande masivo, de estructura interna multilamelar, de forma groseramente hemisférica achatada en los ejemplares de menor talla y generalmente piriforme alargada, aplanada por una de las caras y ligeramente curvada en los ejemplares más grandes. Superficie cubierta con numerosas cámaras de cría desgastadas. La colonia está atravesada en su superficie más aplanada por un conducto (puede haber dos o, menos frecuentemente, más), delatado por uno o dos orificios. El que está siempre presente se ubica generalmente en el extremo más apuntado de la colonia. El segundo orificio ocupa una posición postero-central, en la mitad más ancha de la colonia. A veces, en vez de túnel o túneles hay profundas acanaladuras longitudinales, más o menos curvadas.
Observaciones: La silicificación que presentan estos fósiles (ver apartado de Procesos tafonómicos) hace difícil su identificación. Idealmente, es necesario que los gonozoides estén bien conservados y que se pueda determinar si tienen un techo con pared interior o exterior. Estos ejemplares podrían pertenecer a una de estas dos familias: Lichenoporidae Smitt, 1867 o Densiporidae Borg, 1944, ya que en ambas hay especies con zoarios de crecimiento multilaminar (Taylor, NHM, com. pers.). Se pueden encontrar varios géneros de ciclostomados comparables desde el Cretácico Inferior (Aptiense) hasta nuestros días. Cellulipora d’Orbigny, 1849, que en el Albiense del sur de Inglaterra normalmente se encuentra silicificado, muestra el mismo tipo de huecos en los centros de las subcolonias que los visibles en los fósiles de Urbasa, pero no parece que estos pertenezcan al mencionado género (Taylor, com. pers.). Los fósiles se parecen algo a Reptomulticava alhamensis Reguant, 1993, del Mioceno de Andalucía. Las estructuras tubulares o acanaladuras que presentan los fósiles de Urbasa podrían ser indicativas de colonias que habrían crecido alrededor de una estructura orgánica hoy perdida, quizás el talo de un alga. Es posible que estos fósiles pertenezcan a un nuevo taxón, pero la formalización de una propuesta en este sentido precisa de un estudio más detallado.
Filo Annelida Lamarck, 1809
Clase Polychaeta Grube, 1850
Clado Canalipalpata Rouse & Fauchald, 1997
Clado (Orden) Sabellida Fauchald, 1977
Familia Serpulidae Rafinesque, 1815
Subfamilia Spirorbinae Chamberlin, 1919
Género Rotularia Defrance, 1827
Especie tipo: Rotularia spirulaea (Lamarck, 1818) por designación posterior (Wrigley, 1951)
Rotularia spirulaea (Lamarck, 1818)
(Fig. 4a-c)
|
 Fig. 4.— Fósiles del Eoceno [Bartoniense–Priaboniense (?)] de Navarra de la Colección Ruiz de Gaona. Rotularia spirulaea (Lamarck, 1818), CRG.5, vista apical-dorsal (a), Urbasa 5; CRG.24, vista umbilical (b), Anderatz; CRG.25, vista apical-dorsal (c), Urbasa 1. Serpulidae indet., CRG.26, vistas apical-dorsal (d) y umbilical (e), Ardanatz 2; CRG.27, vistas apical-dorsal (f) y umbilical (g), Ardanatz 2. Ptychocerithium baylei (Tournouër, 1874), CRG.28, vista apertural (h), Ardanatz 3; CRG.29, vista dorsal? (i), Ardanatz 3. Haustator altavillensis (Cossmann y Pissarro, 1900), CRG.30, (j), Ardanatz 3. Haustator cf. imbricatarius conoideus (J. Sowerby, 1814), CRG.31, (k), Ardanatz 3. Pycnodonte brongniarti (Bronn, 1831), CRG.32, valva izquierda, vista externa (l), Anderatz; CRG.33, valva izquierda, vista externa (m), Urbasa 3. Escalas a-c, h-m, 10 mm; d-g, 5 mm. Fig. 4.— Fósiles del Eoceno [Bartoniense–Priaboniense (?)] de Navarra de la Colección Ruiz de Gaona. Rotularia spirulaea (Lamarck, 1818), CRG.5, vista apical-dorsal (a), Urbasa 5; CRG.24, vista umbilical (b), Anderatz; CRG.25, vista apical-dorsal (c), Urbasa 1. Serpulidae indet., CRG.26, vistas apical-dorsal (d) y umbilical (e), Ardanatz 2; CRG.27, vistas apical-dorsal (f) y umbilical (g), Ardanatz 2. Ptychocerithium baylei (Tournouër, 1874), CRG.28, vista apertural (h), Ardanatz 3; CRG.29, vista dorsal? (i), Ardanatz 3. Haustator altavillensis (Cossmann y Pissarro, 1900), CRG.30, (j), Ardanatz 3. Haustator cf. imbricatarius conoideus (J. Sowerby, 1814), CRG.31, (k), Ardanatz 3. Pycnodonte brongniarti (Bronn, 1831), CRG.32, valva izquierda, vista externa (l), Anderatz; CRG.33, valva izquierda, vista externa (m), Urbasa 3. Escalas a-c, h-m, 10 mm; d-g, 5 mm.
|
|
|
 Fig. 5. — Fósiles del Eoceno [Bartoniense–Priaboniense (?)] de Navarra de la Colección Ruiz de Gaona. Pycnodonte rarilamella(Melleville, 1843), CRG.34, valva izquierda, vistas externa (a) e interna (b), Zabaldika. Chlamys biarritzensis (d’Archiac, 1846), CRG.35, valva derecha, vista externa (c), Urbasa 5; CRG.36 (sigla original: 2126), valva derecha, vista externa (d), Anderatz. Spondylus cisalpinus Brongniart, 1823, CRG.37, valva derecha, vista externa (e), valva izquierda, vista externa (f), Anderatz; CRG.38, valva derecha, vista externa (g), Urbasa 2. Dimya pamplonensis (Carez, 1881), CRG.39, valva derecha, vistas externa (h) e interna (i); CRG.40, valva izquierda, vista externa (j), Oritz. Escalas a-g, 10 mm; h-j, 5 mm. Fig. 5. — Fósiles del Eoceno [Bartoniense–Priaboniense (?)] de Navarra de la Colección Ruiz de Gaona. Pycnodonte rarilamella(Melleville, 1843), CRG.34, valva izquierda, vistas externa (a) e interna (b), Zabaldika. Chlamys biarritzensis (d’Archiac, 1846), CRG.35, valva derecha, vista externa (c), Urbasa 5; CRG.36 (sigla original: 2126), valva derecha, vista externa (d), Anderatz. Spondylus cisalpinus Brongniart, 1823, CRG.37, valva derecha, vista externa (e), valva izquierda, vista externa (f), Anderatz; CRG.38, valva derecha, vista externa (g), Urbasa 2. Dimya pamplonensis (Carez, 1881), CRG.39, valva derecha, vistas externa (h) e interna (i); CRG.40, valva izquierda, vista externa (j), Oritz. Escalas a-g, 10 mm; h-j, 5 mm.
|
|
1818 Serpula spirulaea – Lamarck: 366.
Sinonimias completas en Mikuž (2008: 162).
Material: Cuarenta y siete ejemplares provenientes de Anderatz (Abartzutza, Navarra), Formación Urbasa-Andia, Bartoniense. Dieciséis ejemplares de Urbasa 1 (sierra de Urbasa, Navarra), parte superior de la Formación Urbasa-Andia, unidad cartográfica 268, Mapa Geológico de Navarra 1:25:000, Bartoniense. Ciento nueve ejemplares, algunos muy fragmentarios, de Urbasa 4 (sierra de Urbasa, Navarra), unidad cartográfica 267, Mapa Geológico de Navarra 1:25:000, Priaboniense (?). Siete ejemplares de Urbasa 5 (sierra de Urbasa, Navarra), Formación Calizas de Urbasa-Andia, unidad cartográfica 263 del Mapa Geológico de Navarra 1:25.000, Bartoniense. A todos los ejemplares les falta total o parcialmente la porción final desenrollada del tubo o chimenea.
Dimensiones:
| Localidad |
A |
E |
| mín. |
x |
máx. |
n |
mín. |
x |
máx. |
n |
| Anderatz |
5,6 |
11,5 |
18,2 |
47 |
2,3 |
3,3 |
5,9 |
47 |
| Urbasa 1 (km 23) |
7,9 |
15,5 |
23,2 |
15 |
3,4 |
4,9 |
7,0 |
15 |
| Urbasa 4 (km 25) |
7,1 |
11,7 |
17,2 |
85 |
2,3 |
3,1 |
4,3 |
85 |
| Urbasa 5 (km 28) |
12,6 |
14,9 |
17,3 |
7 |
3,1 |
4,0 |
6,3 |
7 |
Descripción: Serpúlido provisto de un tubo carbonatado con un enrollamiento visible generalmente planiespiral. Al inicio o vértice del tubo, que siempre falta en los fósiles y puede corresponder a una corta etapa inicial de vida encostrante, le sigue una porción interna de forma troncocónica de crecimiento helicoespiral que, tras las primeras 2–4 vueltas, pasa a una porción visible, predominantemente planiespiral involuta. La superficie de la espira correspondiente al lado de fijación inicial del tubo es más bien plana, en tanto que la opuesta tiende a ser ligeramente cóncava. El tubo desarrolla una carena o quilla periférica dando lugar a secciones del mismo subtriangulares. La forma troncocónica puede persistir hasta un estado avanzado de crecimiento dando lugar a especímenes de forma cónica (conical specimens, según Savazzi, 1995). Aunque solo se puede observar parcialmente en 5 ejemplares de la localidad de Anderatz y 5 de Urbasa 4, en Rotularia spirulaea la porción final del tubo suele estar desenrollada, formando una estructura en forma de chimenea (chimney, según Seilacher et al., 2008).
La superficie externa del tubo es bastante lisa, marcándose los surcos o líneas radiales de crecimiento —más bien rectas o ligeramente onduladas— correspondientes a los sucesivos depósitos de lamelas. Suele destacar un neto surco o acanaladura espiral dorsal en ambos lados del tubo, que marca la base de la quilla. Las secciones del lumen pasan de almendradas (ovaladas afiladas) en la parte cónica del tubo, a subcirculares afiladas en la porción planiespiral del mismo para, finalmente, en la apertura hacerse circulares. Siguiendo a Savazzi (1995), orientados los ejemplares con el vértice hacia arriba, el sentido de enrollamiento de los especímenes estudiados es mayoritariamente levógiro. Tanto en los fósiles de Anderatz como en los de Urbasa (especialmente en los de Urbasa 4 y 5) estas y otras características del tubo se reconocen de manera incompleta al haber sufrido, durante la diagénesis temprana, una intensa silicificación con formación de anillos de beekita que desfiguran total o parcialmente la morfología originaria (Fig. 4a-b, ver apartado de Procesos tafonómicos).
Observaciones: Rotularia spirulaea (Lamarck, 1818) es una especie común en el Eoceno del dominio del Tetis (Fozy & Szente, 2014; Hoşgör & Okan, 2006; Mikuž, 2008; Savazzi, 1995), incluida la cuenca de Aquitania, en el área norpirenaica (Lamarck, 1818; d’Archiac, 1846, 1850a; Rouault, 1850; Bouillé, 1873, 1876). De esta región proviene el material tipo de la especie, del que se conserva un sintipo en el Muséum national d’Histoire naturelle de Paris, recogido en los alrededores de la ciudad de Baiona-Bayonne (espécimen MNHN.F.R06729, colección Lamarck 2008-14).
R. spirulaea es frecuente en el área surpirenaica. Las primeras noticias sobre su presencia provienen del geólogo francés Carez (1881) quien, precisamente, denominó las Margas de Pamplona, junto con las de Vic (Cataluña) y las de Jaca y la Canal de Berdún (Aragón), como “Marnes bleues à Serpula spirulaea”. Las primeras citas de R. spirulaea en las formaciones margosas de Navarra provienen de trabajos posteriores, en concreto los de Ruiz de Gaona (1947a), Mendizabal & Ruiz de Gaona (1949) y Ruiz de Gaona & Colom (1950). La presencia de R. spirulaea en la zona de este estudio ha sido nuevamente constatada por Astibia et al. (2014, 2016), siendo sus fósiles localmente muy abundantes en niveles margosos de las formaciones Areniscas de Ardanatz y Margas de Ilundain, en asociación con otros muchos macrofósiles (corales, briozoos, moluscos, equinodermos y otros grupos). Dos trabajos recientes de Elorza & Astibia (2017, 2018) estudian en detalle la tafonomía y paleobiología de este taxón en Navarra. La constatación de la presencia de R. spirulaea en los niveles del Bartoniense de las localidades de Anderatz y de la sierra de Urbasa, gracias a los ejemplares de la colección Ruiz de Gaona, suponen extender geográficamente la existencia de este taxón a nuevas zonas del área surpirenaica.
Serpulidae indet.
(Fig. 4d-g)
Material: Nueve ejemplares provenientes de Ardanatz 2 (Eguesibar, Navarra), parte alta de la Formación Margas de Pamplona o parte baja de la Formación Margas de Ilundain), Bartoniense.
Dimensiones: A = 10,0–6,8; H = 3,1–2,2
Descripción: Tubo enrollado de poliqueto encostrante, adherido generalmente a una colonia de briozoos (episkeletozoan, según Taylor & Wilson, 2003). Las vueltas del tubo se solapan parcialmente. Orientado el tubo según el criterio de Jäger (1983) y Savazzi (1995), esto es, con el vértice hacia arriba (oculto por el briozoo sobre el que de adhiere), el enrollamiento en todos los ejemplares es levógiro. Ornamentación externa longitudinal y transversal del tubo muy marcada, con una costilla o cordón dorsal longitudinal aquillado que aumenta de talla junto al peristoma. Base acordonada, claramente hueca en algunos ejemplares, a ambos lados del tubo y apretadas costillas transversales en zig-zag o algo granulosas que se transforman a totalmente granulosas en la base. La sección del tubo es subtriangular, redondeada por los flancos. La sección del lumen es circular.
Observaciones: En la etiqueta original los fósiles figuran como Serpula sp. (Ardanaz, km 2,5). La compleja ornamentación transversal es parecida a la de especies como Proliserpula hemmoorensis Jäger, 1983 o Hamulus sexangularis (Münster en Goldfuss, 1831), del Maastrichtiense superior del noroeste de Alemania (Jäger, 1983), pero las secciones del tubo del serpúlido de Ardanatz, subtriangulares-trapezoidales y el enrollamiento del mismo difieren de las secciones subcirculares y hexagonales, respectivamente, de las especies mencionadas. El tubo enrollado, con vueltas que se solapan y rugosidad lateral, es parecido al de la especie actual Serpula hartmanae Reish, 1968 (ver Ben-Eliahu & Ten Hove, 2011, fig. 28D). Es probable que estos fósiles correspondan a una nueva especie, pero su proposición formal precisa de un estudio más detallado.
Filo Mollusca Linnaeus, 1758
Clase Gastropoda Cuvier, 1795
Clado (Superorden) Caenogastropoda Cox, 1960
Clado (Orden) Sorbeoconcha Ponder & Lindberg, 1997
Superfamilia Cerithioidea Fleming, 1822
Familia Cerithiidae Fleming, 1822
Género Ptychocerithium Sacco, 1895
Especie tipo: Cerithium granulinum Bellardi & Michelotti, 1840 por designación original.
Ptychocerithium baylei (Tournouër, 1874)
(Fig. 4h-i)
1874 Cerithium baylei – Tournouër: 528.
1876 Cerithium sp. – Tournouër, en Bouillé: 250.
1873 Cerithium suessi – Tournouër en Bouillé [non Gemmellaro, 1868]: 463, pl. 5, fig. 12.
1911 Cerithium johannae – Boussac: 34, pl.7, fig. 16.
2016 Ptychocerithium johannae – Astibia et al.: 13, fig. 6a.
|
 Fig. 6.— Fósiles del Eoceno [Bartoniense–Priaboniense (?)] de Navarra de la Colección Ruiz de Gaona. Venericardia hortensis (Vinassa de Regny, 1897), CRG.41, valva izquierda, vista externa (a), valvas izquierda y derecha, vista umbonal (b), Ardanatz 3. Chama granulosa d’Archiac, 1850, CRG.42, vista externa (c), Ardanatz 3; CRG.43, vista externa (d), Ardanatz 3. Harpactoxanthopsiscf.quadrilobatus(Desmarest, 1817), CRG.44 (sigla original: 2338), vistas dorsal (e) y ventral (f), Urbasa 2. Isselicrinuscf.didactylus (d’Orbigny en d’Archiac, 1846), CRG.45, columnar internodal, vista articular (g), Tiebas; CRG.46, columnar nodal, vista articular distal (h), Tiebas; CRG.46, fragmento de pedúnculo con columnar nodal en su extremo inferior, vista lateral (i), Tiebas. Metacrinus sp., CRG.49, columnar internodal proximal, vista articular (j), Ardanatz 2; CRG.50, columnar intermodal, vista articular (k), Ardanatz 2; CRG.51, fragmento de pedúnculo con columnar nodal en su extremo inferior, vista lateral (l), Ardanatz 2. Escalas a-b, g-l, 5 mm; c-f, 10 mm. Fig. 6.— Fósiles del Eoceno [Bartoniense–Priaboniense (?)] de Navarra de la Colección Ruiz de Gaona. Venericardia hortensis (Vinassa de Regny, 1897), CRG.41, valva izquierda, vista externa (a), valvas izquierda y derecha, vista umbonal (b), Ardanatz 3. Chama granulosa d’Archiac, 1850, CRG.42, vista externa (c), Ardanatz 3; CRG.43, vista externa (d), Ardanatz 3. Harpactoxanthopsiscf.quadrilobatus(Desmarest, 1817), CRG.44 (sigla original: 2338), vistas dorsal (e) y ventral (f), Urbasa 2. Isselicrinuscf.didactylus (d’Orbigny en d’Archiac, 1846), CRG.45, columnar internodal, vista articular (g), Tiebas; CRG.46, columnar nodal, vista articular distal (h), Tiebas; CRG.46, fragmento de pedúnculo con columnar nodal en su extremo inferior, vista lateral (i), Tiebas. Metacrinus sp., CRG.49, columnar internodal proximal, vista articular (j), Ardanatz 2; CRG.50, columnar intermodal, vista articular (k), Ardanatz 2; CRG.51, fragmento de pedúnculo con columnar nodal en su extremo inferior, vista lateral (l), Ardanatz 2. Escalas a-b, g-l, 5 mm; c-f, 10 mm.
|
|
Material: Dieciocho ejemplares, algunos en estado muy fragmentario, todos incompletos en la parte apical y la apertura, provenientes de Ardanatz 3 (Eguesibar, Navarra), transición entre las formaciones Areniscas de Ardanatz y Margas de Ilundain, Bartoniense.
Dimensiones: H = (36,8)–(14,4); A = (15,0)–(8,5)
Descripción: Concha ceritiforme multiespiral, con alrededor de 12 vueltas en especímenes adultos; teleoconcha larga, turriculada. La ornamentación espiral consta de tres cordones primarios, nodulosos, bien desarrollados, entre los que se intercalan cordones secundarios perlados muy finos, y cordones terciarios entre estos. En las dos últimas vueltas de la espira y en la última vuelta de la concha el cordón espiral primario más adapical tiende a ser más prominente. Las líneas de crecimiento son opistocirtas-ortoclinas. Los nódulos o tubérculos de los cordones espirales se desarrollan en la intersección con costillas axiales generalmente no muy prominentes. La última vuelta de la concha presenta tres cordones espirales con tubérculos más alargados que en la espira, no menos de ocho cordones o costillas espirales menores y una fuerte variz axial opuesta a la apertura (lado abapertural). En todos los ejemplares disponibles la apertura está rota; no obstante, parece relativamente pequeña, suponiendo quizás del 16 al 20 por ciento de la longitud total de la concha, con aparentes restos de un labio interno bien desarrollado.
Observaciones: La cita más antigua sobre Cerithiidae en el Eoceno de Navarra se debe a Hernández Sampelayo (1933), quien menciona la presencia de las especies Cerithiun pseudocorrugatum y Cerithium lamellosum en los bancos de caliza del flysch de la población de Esa-Yesa. La presencia de Cerithiidae (Cerithium sp.) en el Bartoniense de Ardanatz ya fue mencionada por Ruiz de Gaona (1947a), Mendizábal & Ruiz de Gaona (1949) y Ruiz de Gaona & Colom (1950). Esta especie es la más abundante entre los gasterópodos fósiles de las Areniscas de Ardanatz y ha sido descrita en detalle en Astibia et al. (2018). También está presente, aunque es mucho menos frecuente, en los afloramientos del cercano valle de Itzagaondoa (Formación Margas de Ilundain) (Astibia et al., 2018). Una especie estrechamente relacionada con Ptychocerithium baylei es P. johannae (Tournouër en Bouillé, 1873), del Eoceno norpirenaico de la Formación Margas de la Côte des Basques, en Biarritz (Bartoniense–Priaboniense, cuenca de Aquitania; Jacquot, 1864; Mathelin & Sztrakos, 1993). Sin embargo, en P. johannae las granulaciones de los cordones espirales primarios solo están bien desarrolladas en las primeras vueltas de la teleoconcha, atenuándose significativamente en las siguientes, dando como resultado cordones espirales más suaves que en P. baylei. La morfología también es bastante similar a la de Cerithium biarritzense (ver Bouillé, 1876: 62, pl. 3, fig. 5), otra especie de los “Cérites des couches à Serpula spirulaea” (en denominación de Boussac 1911) de las margas de la Côte des Basques. Es probable que P. baylei y C. biarritzense sean especies sinónimas (Astibia et al., 2018).
Familia Turritellidae Lovén, 1847
Género Haustator Montfort, 1810
Especie tipo: Haustator gallicus Montfort, 1810 por monotipia.
Haustator altavillensis (Cossmann & Pissarro, 1900)
(Fig. 4j)
1900 Turritella altavillensis – Cossmann & Pissarro: 196, pl. 20, figs 6–7.
1956 Turritella (Haustator) altavillensis – Villalta Comella: 143–144, pl. 3, figs 1a–1e.
Material: Un ejemplar incompleto, con la concha parcialmente disuelta, sin la porción apical ni la apertura, proveniente de Ardanatz 3 (Eguesibar, Navarra), transición entre las formaciones Areniscas de Ardanatz y Margas de Ilundain, Bartoniense.
Descripción: Aunque el ejemplar estudiado es muy fragmentario y no está bien conservado, se observan en el mismo algunos de los rasgos morfológicos de esta especie: concha turriculada con vueltas de lados rectilíneos, basalmente carenadas, ornamentadas con cordones o costillas espirales afiladas, más apretadas en la mitad adapical, más separadas y desarrolladas en la mitad abapical de cada vuelta. Cerca de la base de cada vuelta se desarrolla una costilla espiral más prominente.
Observaciones: La cita más antigua sobre Turritellidae en el Eoceno de Navarra se debe a Hernández Sampelayo (1933), quien menciona la presencia de fósiles del género Turritella en los bancos de caliza del flysch de Esa-Yesa. Más tarde, Ruiz de Gaona (1947a), Mendizábal & Ruiz de Gaona (1949) y Ruiz de Gaona & Colom (1950) indican la presencia de Turritellidae entre los gasterópodos de las formaciones margosas del Eoceno de la cuenca de Pamplona. Ruiz de Gaona & Colom (1950) citan la especie Turritella duvali en el Bartoniense de Ardanatz-Eguesibar. Haustator altavillensis ha sido descrita en detalle por Astibia et al. (2018), a partir de ejemplares provenientes de las secciones de Ardanatz-Eguesibar y del cercano valle de Itzagaondoa.
La morfología general de este taxón es bastante similar a la de Turritella asperula Brongniart, 1823 del Paleógeno de la Chambre d’Amour en Biarritz (cuenca de Aquitania), descrita y figurada por Boussac (1911: 82–83, pl. 21, fig. 15 y pl. 22, figs. 3-4), pero la disposición de las costillas o cordones espirales es diferente. Tanto en el ejemplar de la CRG como en los descritos por Astibia et al. (2018), las costillas son más fuertes y más separadas en la parte abapical de cada vuelta, mientras que en Haustator asperulus se distribuyen más regularmente en toda la superficie de cada vuelta. Por el contrario, los fósiles de Navarra son muy similares a los que Villalta Comella (1956: 143–144, pl. 3, figs. 1a-1e) asigna a la especie Turritella (Haustator) altavillensis Cossmann & Pissarro (1900), del Bartoniense del Valle de Basa / Ballibasa (Huesca, cuenca de Jaca).
Haustator cf. imbricatarius conoideus (J. Sowerby, 1814)
(Fig. 4k)
1814 Turritella conoidea – Sowerby J.: 109, pl. 51, figs 1–2, 4.
Material: Un ejemplar incompleto, con la concha parcialmente disuelta, sin la parte apical ni la apertura, proveniente de Ardanatz 3 (Eguesibar, Navarra), transición entre las formaciones Areniscas de Ardanatz y Margas de Ilundain, Bartoniense.
Descripción:Concha turriculada, pronunciadamente cónica; teleoconcha con suturas profundas y vueltas espirales bien individualizadas, de perfiles laterales ligeramente convexos y ornamentación formada por cordones espirales de al menos tres órdenes, finamente granulosos y regularmente espaciados.
Observaciones: Fósiles similares provenientes de Ardanatz-Eguesibar y de los afloramientos del valle de Itzagaondoa han sido descritos y discutidos en detalle por Astibia et al. (2018). Haustator imbricatarius (Lamarck, 1804) es una especie que está presente en las cuencas de París y Londres, entre otras áreas (ver Courville et al., 2012). D’Archiac (1846) y Boussac (1911) citaron la presencia de la especie Turritella imbricataria Lamarck, 1804 del Eoceno de Biarritz, en la parte sur de la Côte des Basques (cuenca de Aquitania). Como indican Astibia et al. (2018), los fósiles de Navarra tienen una mayor similitud con varios especímenes de Haustator imbricatarius conoideus (J. Sowerby, 1814) de Barton-on-Sea (Inglaterra) incluidos en la colección Cossmann del MNHN de París. Haustator elongatus (J. Sowerby, 1814) del Bartoniense de Barton-on-Sea también tiene vueltas de aspecto imbricado y una silueta algo convexa, pero la concha es más estrecha, con un ángulo apical más pequeño que en H. imbricatarius conoideus y en los fósiles de Navarra.
Clase Bivalvia Linnaeus, 1758
Infraclase Pteriomorphia Beurlen, 1944
Orden Ostreida Férussac, 1822
Suborden Ostreidina Férussac, 1822
Superfamilia Ostreoidea Rafinesque, 1815
Familia Gryphaeidae Vialov, 1936
Subfamilia Pycnodonteinae Stenzel, 1959
Género Pycnodonte Fischer de Waldheim, 1835
Especie tipo: Pycnodonte radiata Fischer de Waldheim, 1835 por designación original.
Pycnodonte brongniarti (Bronn, 1831)
(Fig. 4l-m)
1831 Gryphaea brongniarti – Bronn: 630.
Sinonimias completas en Abad (2001: 548).
Material: Cuatro ejemplares incompletos provenientes de Anderatz (Abartzutza, Navarra), Formación Calizas de Urbasa-Andia, Bartoniense. Tres ejemplares incompletos de Urbasa 3 (sierra de Urbasa, Navarra), unidad cartográfica 267, Mapa Geológico de Navarra 1:25:000, Priaboniense (?).
Dimensiones:(Valva izquierda) H = (40,0)–(25,7); L = (28,4)–(18,0)
Descripción: Concha grifeiforme (sensu Carter et al., 2012) de talla media, fuertemente inequivalva, inequilateral e irregular, más alta (larga) que ancha. Valva izquierda fuertemente convexa, arqueada, con umbo prosogiro, prominente y recurvado. Los bordes laterales son rectos y divergentes. Un surco posterior radial que se origina debajo del área umbonal lleva al desarrollo de un ala o lóbulo convexo, que puede proyectarse hacia atrás de manera prominente. La superficie externa de la concha es bastante lisa, con líneas de crecimiento bajas e irregulares. Debido a lo fragmentario y al duro relleno sedimentario de los fósiles disponibles no es posible describir la morfología de la charnela, ni de la impresión muscular, en la parte interna de las valvas. No hay en la CRG ninguna valva derecha de este taxón.
Observaciones: Los ejemplares de Anderatz figuran etiquetados en la CRG como Pycnodonta archiaci y los fósiles provenientes de Urbasa, más fragmentarios, como Pycnodonta sp. Abad (2001) considera las especies P. archiaci (Bellardi, 1852) y P. brongniarti (Bronn, 1931) como sinónimas. P. brongniarti es frecuente en los afloramientos de la parte alta de la Formación Margas de Ilundain, en el sinclinal de Aranguren-Itzaga, cuyos fósiles se describen en detalle en Astibia et al. (2018). Una especie eocénica cercana a P. brongniarti es Pycnodonte pharaonum (Oppenheim, 1903). Sin embargo, en P. pharaonum la valva izquierda es más convexa y adquiere en la parte media una forma aguda con perfil de quilla, de donde pueden partir una o dos espinas hióticas, ausentes en otras species del mismo género (Abad, 2001). P. brongniarti tiene una amplia distribución bioestatigráfica (Paleoceno–Mioceno) y paleobiogeográfica (norte de África y el sur de Eurasia). Es abundante en numerosas localidades del Eoceno (Bartoniense–Priaboniense) de las regiones catalanas de Igualada y Vic, en la parte oriental de la cuenca del Ebro. P. brongniarti también está presente en varios afloramientos de los alrededores de Angelu-Anglet y Biarritz, en la parte alta de la serie del Paleógeno (? Oligoceno inferior, Rupeliense) de la costa vasca (cuenca de Aquitania), denominada como Ostrea vesicularis por d’Archiac (1846, 1850a), Ostrea brongniarti y Ostrea vesiculosa Sowerby var. nummulítica por Tournouër en Bouillé (1873, 1876) y Ostrea brongniarti por Boussac (1908, 1911). Cossmann (1921) también describió fósiles de Liostrea (Pycnodonta) brongniarti de los mismos afloramientos de la cuenca de Aquitania. Siguiendo la descripción de Abad (2001), tanto Ostrea vesicularis como O. vesiculosa var. nummulitica encajarían dentro del rango de variabilidad de la especie P. brongniarti.
Pycnodonte rarilamella(Melleville, 1843)
(Fig. 5a-b)
1843 Ostrea rarilamella – Melleville: 42–43.
1860 Ostrea rarilamella – Deshayes: tomo 1, láms. 81, 82.
Material: Una valva izquierda incompleta proveniente de Zabaldika (Esteribar, Navarra), parte superior del Miembro Calciclástico 3 de la Formación Anotz, en su transición lateral a la Formación Erro del Grupo Hecho, Luteciense medio.
Dimensiones: H = (107,3); L = > (103,2); E = (52,0).
Descripción: Valva izquierda de tamaño mediano-grande, de contorno sub-redondeado, irregular, algo más alta que ancha; gruesa y fuertemente convexa, con un surco radial que origina una pequeña ala o lóbulo posterior. Superficie externa lisa, ligeramente ondulada en sentido umbo-paleal, con fina y apretada estriación de crecimiento, con algunos ribetes más destacados, irregularmente espaciados. El interior de la valva es liso y poco profundo. El área ligamentaria se conserva y es pequeña, de contorno subtriangular (?), con numerosas estrías bien visibles. El resilifer es curvo y cóncavo. Debajo del área ligamentaria se observa una pequeña depresión correspondiente a la inserción del músculo de Quenstedt. Presenta una neta impresión ovalada del músculo aductor, ubicada en el cuadrante supero-posterior de la valva.
Observaciones: La talla y morfología de este ejemplar se corresponden en términos generales con las de las especies Pycnodonte gigantica (Solander en Brander, 1766) y Pycnodonte rarilamella (Melleville, 1843), presentes en el Eoceno y Oligoceno de Europa y Asia, incluido el dominio pirenaico (De Bouillé, 1873; Boussac, 1911; Martinius, 1991; Abad, 2001). Según Abad (2001: 563), P. rarilamella se distingue fácilmente de P. gigantica debido a que la primera tiene una valva izquierda con una forma más aguda y aquillada, a menudo con un surco radial que diferencia una zona posterior, mientras que P. gigantica presenta una forma convexa más suave o laxa. Esta morfología acercaría más el fósil de la CRG a la especie P. rarilamella. La descripción original de la especie Ostrea rarilamella realizada por Melleville (1843) no incluye figuración alguna. Por otro lado, Deshayes (1860, pls. 81, 82), en su descripción de los fósiles de los alrededores de París, figura una valva izquierda atribuida a esta especie —a grandes rasgos similar al ejemplar de Zabaldika— que, además de las ondulaciones y estrías de crecimiento paralelas al contorno de la concha, muestra una superficie externa con marcados pliegues o costillas radiales, prácticamente ausentes en el ejemplar navarro. Aunque algo más atenuados, dichos pliegues o costillas radiales también son claramente visibles en un ejemplar atribuido a P. rarilamella figurado por Boschele et al. (2016), proveniente del Oligoceno inferior (Rupeliense inferior) de Valsugana (Trentino, Italia). De lo dicho, cabe albergar dudas sobre la asignación aquí planteada; sin embargo, en la descripción original de la especie Melleville (1843: 43) viene a indicar que dicha ondulación radial no se da sino a veces (“A l’extérieur elle [la valva izquierda] porte quelques lames transversales, irrégulières, très-minces, écailleuses et presque couchées, quelquefois plissées d’une manière peu sensible”). Esta matización en la diagnosis original de P. rarilamella permitiría fundamentar la clasificación aquí propuesta.
Otra especie que puede presentar una morfología parecida es Hyotissa martinsii (d’Archiac, 1850a). Sin embargo, tanto el fósil estudiado como P. rarilamella y P. gigantica pueden diferenciarse de H. martinsii en que en la valva izquierda no se desarrollan costillas, normalmente presentes y fuertes en la última especie.
Orden Pectinida J. Gray, 1854
Suborden Pectinida J. Gray, 1854
Superfamilia Pectinoidea Rafinesque, 1815
Familia Pectinidae Rafinesque, 1815
Género Chlamys Röding, 1798
Especie tipo: Pecten islandicus Müller, 1776 por designación posterior (Herrmannsen, 1846).
Chlamys biarritzensis (d’Archiac, 1846)
(Fig. 5c-d)
1846 Pecten biarritzensis – d’Archiac: 210–211, pl. 8, fig. 9a, b.
Sinonimias completas en Abad (2001: 419–420).
Material: Seis valvas algo incompletas, entre las que hay probablemente dos derechas y una izquierda, provenientes de Anderatz (Abartzutza, Navarra), Formación Calizas de Urbasa-Andia, Bartoniense. Una valva derecha (?) de Urbasa 5 (sierra de Urbasa, Navarra), Formación Calizas de Urbasa-Andia, unidad cartográfica 263 del Mapa Geológico de Navarra 1:25.000, Bartoniense. Todos los ejemplares están incompletos dorsalmente.
Dimensiones: Urbasa 5, valva derecha (?) H = 38,8; L = 36,8 / Anderatz, valva derecha (?) H = 35,9–31,5; L = 33,0–29,0; valva izquierda (?) H = 31,9–(21,0); L = (29,5)–19,3
Descripción: Concha con contorno ventral oval, ligeramente más alta que ancha, con márgenes laterales ligeramente curvados. La ornamentación está compuesta por al menos 19–21 costillas radiales que presentan una forma estriada o lobulada longitudinalmente, sobre todo en su tercio final, siendo el lóbulo central el más desarrollado. Los espacios intercostales son ligeramente más estrechos que las costillas. La superficie externa de las costillas está recorrida longitudinalmente por dos, tres o más finos cordones o crestas, escamosas o dentadas, con lóbulos apuntando hacia el borde ventral de la concha, resultado de la intersección de una apretada sucesión de estrías transversales onduladas y de aspecto imbricado. Cada valva posee dos orejetas, anterior y posterior, conservadas parcialmente en cuatro de los ejemplares estudiados. Las orejetas están externamente ornamentadas con una serie de finas cóstulas escamosas, que van desde el umbo hasta el borde de las mismas. El borde de la orejeta anterior se conserva en uno de los ejemplares de Anderatz y presenta la entrada del seno bisal. El borde de la orejeta posterior es más rectilíneo.
Observaciones: Ruiz de Gaona & Colom (1950) indican la abundancia de “Pecten subtripartitus” en los niveles areniscosos de Ardanatz-Eguesibar, formándose en algunos casos auténticas lumaquelas. Efectivamente, tal sería el caso observado por Astibia et al. (2018, Fig. 3A) en la sección AD3 de la mencionada localidad. No obstante, a día de hoy no existen ejemplares de este taxón en la CRG que provengan de Ardanatz.
La morfología aquí descrita corresponde al ejemplar de Urbasa. En los fósiles de Anderatz su intensa silicificación ha modificado la ornamentación original de la concha (Fig. 5d, ver apartado de Procesos tafonómicos). Las características descritas son similares a las de las especies Chlamys biarritzensis (d’Archiac, 1846) y Chlamys subtripartita (d’Archiac, 1850) del Eoceno “nummulítico” de Biarritz (Cuenca de Aquitania), inicialmente asignadas al género Pecten (“Pecten subtripartitus”). Estas especies también fueron descritas y figuradas por Boussac (1911) y Cossmann (1921). D’Archiac (1950) creó varias especies más, que Tournouër en Bouillé (1876) sugirió, por el contrario, podían considerarse sinónimas de “Pecten biarritzensis”. Boussac (1911) indicó que la ornamentación es más marcada y escamosa en C. biarritzensis que en C. subtripatita, y que ambas especies parecían estar separadas estratigráficamente. Sin embargo, Oppenheim (1901), Fabiani (1915) y Piccoli & Mocellin (1962) consideraron su sinonimia. En esta misma línea de opinión, Abad (2001) estudió numerosos fósiles del Eoceno de Cataluña (Igualada y Manresa en Barcelona y Vic en Girona), que asignó en su totalidad a una única especie, C. biarritzensis, asimismo presente en muchos otros lugares del Eoceno (Luteciense–Priaboniense) y Oligoceno de Europa y África del Norte (ver Abad, 2001).
Familia Spondylidae J. Gray, 1826
Género Spondylus Linnaeus, 1758
Especie tipo: Spondylus gaederopus Linnaeus, 1758 por designación posterior (Schmidt, 1818).
Spondylus cisalpinus Brongniart, 1823
(Fig. 5e-g)
1823 Spondylus cisalpinus – Brongniart: 76, pl. 5, fig. 1a–c.
Material: Un ejemplar algo deteriorado en la región umbonal proveniente de Anderatz (Abartzuza, Navarra), Formación Calizas de Urbasa-Andia, Bartoniense. Un ejemplar proveniente de Urbasa 2 (sierra de Urbasa, Navarra), zona límite entre las unidades cartográficas 268 (Formación Urbasa-Andia) y 267, Mapa Geológico de Navarra 1:25:000, Bartoniense–Priaboniense (?). Un fragmento de valva derecha proveniente de Ardanatz 3 (Eguesibar, Navarra), transición entre las formaciones Areniscas de Ardanatz y Margas de Ilundain, Bartoniense.
Dimensiones: Anderatz, valva derecha H = 39,6; L = 42,2; E = 26,0 / Urbasa 2, valva derecha H = 56,9; L = 62,0; E = 33,2
Descripción: Ejemplar de Anderatz (CRG.37): Concha inequivalva, inequilateral, de contorno ovalado. La valva derecha es fuertemente convexa, con una pequeña superficie de fijación aplanada de contorno subcircular. Sus bordes laterales forman un ángulo de unos 104 grados. La superficie externa está ornamentada con unas 12–14 costillas radiales primarias lisas, de sección redondeada, provistas de numerosas espinas regularmente dispuestas en intervalos cortos a lo largo de las mismas. Superficies intercostales desarrolladas, bastante planas, rugosas, donde quedan patentes las líneas de crecimiento y una serie de cóstulas —aproximadamente de 2 a 7— finas y escamosas, dentadas, que en la parte central del espacio intercostal pueden desarrollarse más dando lugar a costillas radiales secundarias. La valva izquierda es más aplanada, menos convexa y de tamaño, en sentido umbopaleal, ligeramente menor que la valva derecha. En el ejemplar descrito apenas puede observarse su morfología externa, debido a que se halla cubierta por sedimento fuertemente litificado. Como ya se ha indicado en relación con los fósiles de Anderatz, el fósil está silicificado, cubierto por conjuntos de anillos de beekita que han alterado la morfología originaria de la concha (Fig. 5e, ver apartado de Procesos tafonómicos).
Ejemplar de Urbasa 2 (CRG.38): Concha inequivalva e inequilateral, de contorno ovalado. Los bordes anterior y posterior de la concha forman en la región umbonal un ángulo abierto de 90º o más. La valva derecha es muy convexa con su mitad superior aplanada, en parte probablemente por distorsión fosildiagenética (sensu Fernández López, 2000), de forma que la sección anteroposterior es de contorno trapezoidal. La región umbonal muestra una pequeña área provista de ornamentación lamelar comarginal. El resto de la superficie externa exhibe unas 11 costillas radiales primarias lisas, provistas de espinas de sección alta, algo comprimidas lateralmente, dispuestas en sucesión bastante regular a lo largo de las mismas. En las superficies intercostales pueden diferenciarse una o dos costillas secundarias y un número mayor de finas cóstulas dentadas. La valva izquierda es menos cóncava que la valva derecha y con espinas más finas. Está provista de unas 10 costillas radiales y anchos espacios intercostales, con 2 o 3 costillas secundarias y aún más finas cóstulas terciarias intercaladas.
Ejemplar de Ardanatz 3: Se trata de un pequeño fragmento de la región umbonal de una valva derecha, ornamentado con numerosas espinas, regularmente dispuestas sobre las costillas radiales.
Observaciones: La presencia del género Spondylus en el Eoceno surpirenaico de Navarra ha sido mencionada repetidamente por Ruiz de Gaona (1947a), Mendizábal & Ruiz de Gaona (1949) y Ruiz de Gaona & Colom (1950), quienes citan la especie Spondylus eocenus Leymerie, 1881 en el Bartoniense de Ardanatz-Eguesibar. Astibia et al. (2016, 2018) describen y figuran ejemplares provenientes de esa misma localidad y de otras áreas del sinclinal Aranguren-Itzaga, que atribuyen a las especies Spondylus cf. caldesensis Carez 1881, Spondylus cisalpinus Brongniart, 1823 (ecomorfo bifrons), Spondylus cisalpinus Brongniart, 1823 y Spondylus planicostatus d’Archiac, 1847. Spondylus no se había citado hasta el momento en el Eoceno del área de Andia y Urbasa.
El género Spondylus se conoce desde antiguo en el “nummulítico” de Biarritz, en la costa vasca labortana (cuenca de Aquitania), donde fue definida la especie S. planicostatus y donde se han descrito hasta 12 especies diferentes (d’Archiac, 1846, 1850a; Rouault, 1850; Bouillé, 1876; Boussac, 1908, 1911; Cossmann, 1921). La más frecuentemente mencionada es Spondylus subspinosus, que tanto Boussac (1911) como Abad (2001) consideran especie sinónima de Spondylus buchi Philippi, 1846. Spondylus es también muy frecuente en el Eoceno de Cataluña, de donde proviene el holotipo de S. caldesensis (Carez, 1881; Carrasco, 1994; Abad, 2001).
En las etiquetas originales de la CRG los ejemplares aquí descritos figuran como Spondylus eocenus. Pero esta especie, al igual que S. buchi y S. planicostatus —todas ellas incluidas por Zavarei (1973) en un mismo grupo evolutivo (“Grupo de Spondylus buchi”)— presentan conchas muy regulares, equivalvas o sub-equivalvas y equilaterales o sub-equilaterales, diferentes, al menos, de los ejemplares de Anderatz y Urbasa, que exhiben conchas marcadamente inequivalvas e inequilaterales. Asimismo, según las descripciones de Carrasco (1994) y Abad (2001), y a diferencia de los ejemplares aquí descritos, la concha de S. buchi tiene espacios intercostales cortos y angulares. Spondylus planicostatus, taxón definido en el Paleógeno de Biarritz (d’Archiac, 1847, 1850a) presenta costillas más aplanadas y espacios intercostales más apretados que los fósiles de la CRG.
La razón de haber descrito separadamente los ejemplares de la CRG, radica, sobre todo, en que su ornamentación externa es algo diferente; más acusada y espinosa en los de Anderatz y Ardanatz que en el de Urbasa. Pero, según Abad (2001), la ornamentación en S. cisalpinus es muy variable. Por otro lado, el ejemplar de Urbasa parece estar algo más desgastado que los otros dos. Aunque de menor talla, la valva derecha del ejemplar de Anderatz y también el fragmento conservado de Ardanatz presentan una morfología comparable a la del espécimen AG1.4.4 de la localidad navarra de Aranguren, que Astibia et al. (2018: 244, fig. 12C, D) atribuyen a la especie S. cisalpinus (ecomorfo bifrons). Las espinas más numerosas, que se distribuyen regularmente a intervalos cortos en las costillas, las superficies de las costillas más suaves y quizás los espacios intercostales más estrechos, son características que permiten diferenciar estos dos últimos ejemplares y también el de Urbasa 2 de la especie S. caldesensis, que tiene costillas escamosas, con espinas más fuertes e irregularmente distribuidas y espacios intercostales más anchos.
Zavarei (1973) reúne las especies de Spondylus del Paleógeno en tres grupos: el grupo de S. buchi, arriba mencionado, el grupo de S. radula y el grupo de S. cisalpinus, que incluiría además de la especie homónima, a S. bifrons Münster en Goldfuss, 1840 y S. nysti d’Archiac, 1846. Según Carrasco (1994) S. cisalpinus y S. bifrons se separarían por la diferente convexidad de la valva izquierda, mucho menor en la primera especie, y por la ornamentación intercostal de la valva derecha, granulosa en S. cisalpinus y casi lisa en S. bifrons. Estas características acercarían más los ejemplares de la CRG a S. cisalpinus. Sin embargo, para Abad (MGS, com. per., en Astibia et al., 2018) S. bifrons es un ecomorfo de S. cisalpinus, que incluye a los ejemplares que viviendo sobre fondos fangosos desarrollan espinas más fuertes.
En cuanto a S. nysti, especie definida en el Eoceno de Biarritz, la misma se caracteriza sobre todo por presentar una amplia superficie plana en la valva derecha, con una ornamentación formada por lamelas apretadas y paralelas al borde exterior de la concha (comarginales)
(d’Archiac, 1846: 212, Pl. 9, figs. 3–5). Sin embargo, Abad (2001) cuestiona seriamente la validez de esta especie, ya que observa ejemplares de S. cisalpinus con valvas derechas que desarrollan en extensión variable la misma ornamentación lamelar comarginal. Astibia et al. (2018) observan una similar variabilidad en varios ejemplares que asignan a S. cisalpinus de la sección del Bartoniense del valle de Itzagaondoa.
Familia Dimyidae P. Fischer, 1886
Género Dimya Rouault, 1850
Especie tipo: Dimya deshayesiana Rouault, 1850 por monotipia.
Dimya pamplonensis (Carez, 1881)
(Fig. 5h-j)
1881 Plicatula pamplonensis – Carez: 310, pl. 8, figs 2–5.
1911 Dimya richei – Doncieux: 34, pl. 6, figs 7 a–b.
1950 Anomiya pamplonensis – Ruiz de Gaona & Colom: 332, lám. 54, figs 10–13.
Material: Dieciocho ejemplares completos y dos valvas derechas, provenientes de Oritz (Elortzibar, Navarra), parte baja de la Formación Margas de Ilundain, Bartoniense.
Dimensiones: H = 10,8–6,4; L = 9,1–5,3; E = 3,7–2,3
Descripción: Concha pequeña, inequivalva e inequilateral, de contorno ovalado, a veces serrado o irregular, con perfil plano convexo, con el máximo grosor en la región umbonal. El ángulo umbonal varía entre 120º y 160º. La valva izquierda es ligeramente convexa. Su superficie exterior es foliácea con 5 o 6 (7) láminas comarginales, algo onduladas, irregularmente espaciadas y con costillas antimarginales de desarrollo variable, en general, casi imperceptibles. La valva derecha es casi plana, también lamelosa, con costillas muy difuminadas. El extremo umbonal o de fijación está más inclinado que el resto de la valva. El interior de las valvas es liso y aporcelanado. Se pueden distinguir tres regiones o áreas internas: (1) umbonal, correspondiente a la concha interna aragonítica en las especies vivas de Dimya (Waller, 2012), (2) paleal interna y (3) paleal externa, las dos últimas correspondientes al borde calcítico de las especies actuales de Dimya y separadas por una línea de denticulación muy visible (chomata). El área umbonal es profunda, de contorno casi circular, afilada en el extremo superior, separada de la región paleal por una línea curva (línea paleal) de color diferente. Las impresiones de los músculos aductores son generalmente difíciles de observar, la anterior de forma piriforme o triangular en la valva derecha, la posterior bilobulada, con dos componentes ovalados o casi circulares, ubicados en la parte inclinada de la concha. El área paleal interna forma un área o segmento circular, en contacto con el área umbonal, con dos concavidades simétricas para el ligamento, limitadas axialmente por dientes pequeños, o crura, más fuertes en sentido paleal.
Observaciones: La especie Dimya pamplonensis (Carez, 1881) fue descrita a partir de ejemplares fósiles recolectados en niveles margosos al sur de la ciudad de Pamplona (Navarra), depósitos que Carez (1881) consideró equivalentes a las “Margas con Serpula spirulaea” (“Marnes bleues à Serpula spirulaea”) de Cataluña (Igualada, Vic) y Aragón (La Peña, Jaca, Fiscal). Ruiz de Gaona (1947a) y Mendizábal & Ruiz de Gaona (1949) citaron la presencia de esta especie, que asignaron al género Plicatula, en las localidades de Oritz, Altos de Badoztain, Ardanatz-Eguesibar y Zabaltza. Más tarde, Ruiz de Gaona & Colom (1950) asignaron esta especie, cuya presencia amplían a los términos Tejería de Pamplona y Ezkoriz, al género Anomia (“Anomiya”). Calzada & Astibia (1996) incluyeron esta especie en el género Dimya, ya que las valvas muestran dos impresiones musculares y no son perforadas. En el mismo sentido, la atribución genérica a Dimya también fue considerada antes, aunque no publicada, por Ruiz de Gaona, ya que los ejemplares que se guardan en su colección lo hacen con una etiqueta de su puño y letra que pone Dimya pamplonensis Carez. Recientemente, Astibia et al. (2018) han descrito nuevos fósiles de esta especie provenientes de los alrededores de Ardanatz y otros afloramientos de la zona.
Hasta el momento la presencia de D. pamplonensis solo ha sido constatada en las cercanías de Pamplona y zonas colindantes del sinclinal de Aranguren-Itzaga, estando presente en los tramos inferior y medio de las Areniscas de Ardanatz (?) y de las Margas de Ilundain, que representan la sedimentación en fondos blandos y probablemente de mayor profundidad, estando ausente en los tramos altos de estas formaciones donde, por el contrario, es abundante el serpúlido Rotularia spirulaea (Calzada & Astibia, 1996).
D. pamplonensis difiere de Dimya intustriata (d’Archiac, 1850a) del Eoceno de Trabay y Hontet (cuenca de Aquitania, área norpirenaica) por su menor tamaño y por su menor desarrollo de costillas radiales o antimarginales, significativamente más pronunciadas en la segunda especie (Calzada & Astibia, 1996; Astibia et al., 2018).
D. pamplonensis ha sido citada en el Eoceno de Vic (Cataluña) (Farrés & Staid-Staadt 1964) —sin descripción ni figuración— y Sobrarbe (Aragón) (http://fosilesdesobrarbe.blogspot.com.es). En este último caso, cuatro de los ejemplares ilustrados son ligeramente más grandes y tienen costillas más pronunciadas que los fósiles de Pamplona, de forma que podrían asignarse más aceptablemente a Dimya intustriata (d’Archiac, 1850). Por otro lado, la especie Dimya deshayesiana Rouault, 1850 no tiene ornamentación lamelar, estando completamente cubierta con costillas antimarginales (¿radiales?) (Calzada & Astibia, 1996). Waller (2012) incluye D. pamplonensis dentro de las especies válidas de la familia Dimyidae, sinonimizándo Dymia richei Doncieux, 1911 con D. pamplonensis.
Infraclase Heteroconchia Hertwing, 1895
Orden Carditida Dall, 1889
Superfamilia Crasatelloidea Férussac, 1822
Familia Carditidae Férussac, 1822
Subfamilia Venericardiinae Chavan, 1969
Género Venericardia Lamarck, 1801
Especie tipo: Venus imbricata Lamarck, 1801 por designación posterior (Schmidt, 1818).
Venericardia hortensis (Vinassa de Regny, 1897)
(Fig. 6a-b)
1897 Cardita hortensis – Vinassa de Regny: 183–184, pl. 20, fig 4-6.2016 Venericardia junctinoda – Astibia et al.: 15, fig. 6u.
Sinonimias completas en Cossmann (1921).
Material: Un ejemplar de Ardanatz 3 (Eguesibar, Navarra), transición entre las formaciones Areniscas de Ardanatz y Margas de Ilundain, Bartoniense.
Dimensiones: H = 10,1; L = 7,7; E = 5,1
Descripción: Concha equivalva e inequilateral, inflada dorsalmente, de contorno ovalado, acorazonado, con umbos prosogiros prominentes y lúnula corta y profunda. La ornamentación externa de cada valva consiste en (23) 24 costillas radiales fuertes, erizadas con laminillas transversales gruesas, de apariencia escamosa. Los espacios intercostales son profundos, más anchos que las costillas en el tercio anterior de la concha.
Observaciones: Astibia et al. (2018) describen fósiles de esta especie provenientes de la parte alta de la Formación Margas de Ilundain (Navarra). Se trata en general de ejemplares de mayor talla y algo más elongados anteroposteriormente que el descrito en este trabajo. Venericardia hortensis (Vinassa de Regny, 1897) fue definida en el Eoceno de Possagno (Alpes del sur, norte de Italia). También se encuentra en el Eoceno (Bartoniense y Priaboniense) de la Côte des Basques (Biarritz, cuenca de Aquitania), descrita y figurada por Boussac (1911: 44–45) y Cossmann (1921: 124; MNHN.F.J07469). En principio, V. hortensis se puede distinguir de Venericardia junctinoda Cossmann, 1898, del “Nummulítico medio” (Ypresiense) de Àger (cuenca de Àger, Cataluña, área surpirenaica) (Cossmann, 1898; De Renzi, 1971) por su menor tamaño y menor número de costillas radiales (23 frente a 25), pero estas diferencias podrían acomodarse dentro de la variación intraespecífica de una sola especie. Venericardia basteroti (Deshayes, 1851), del Oligoceno (Rupeliense) de Larrat (Landas, cuenca de Aquitania; MNHN.F.A42505), tiene menos costillas que V. hortensis (17 según Cossmann, 1921). Venericardia acuticostata Lamarck, 1806, del Eoceno de Ferme de l’Aulnaie (Oise; MNHN.F.A25073), tiene laminillas más afiladas y estas se desarrollan solo en la parte central de las costillas.
Orden Cardiida Férussac, 1822
Suborden Cardiidina Férussac, 1822
Superfamilia Chamoidea Lamarck, 1809
Familia Chamidae Lamarck, 1809
Género Chama Linnaeus, 1758
Especie tipo: Chama lazarus Linnaeus, 1758 por designación posterior (Children, 1823).
Chama granulosa d’Archiac, 1850
(Fig. 6c-d)
1850 Chama granulosa – d’Archiac: 433, pl. 12, figs 9, 10.
Material: Trece ejemplares incompletos de Ardanatz 3 (Eguesibar, Navarra), transición entre las formaciones Areniscas de Ardanatz y Margas de Ilundain, Bartoniense.
Dimensiones: H = 30,9–(12,2); L = (28,0)–(20,1)
Descripción y observaciones: Chama granulosa es un bivalvo fósil relativamente frecuente en las secciones AD2 y AD4 de Ardanatz-Eguesibar, y ha sido recientemente descrito por Astibia et al. (2018).
El bajo número de laminillas comarginales, su superficie finamente rugosa y sus costillas radiales muy delgadas diferencian estos fósiles de los asignados a Chama pellati Boussac, 1911 del Eoceno (Bartoniense–Priaboniense) de la Côte des Basques (Biarritz, cuenca de Aquitania) y de la cuenca de Pamplona (Astibia et al., 2018). El bajo número de laminillas y la finura de las costillas radiales acercan estos especímenes a la especie C. granulosa D’Archiac, 1850a, especialmente a los especímenes de la Côte des Basques y les Bains figurados por Boussac (1911) y Cossmann (1921).
Filo Arthropoda von Siebold, 1848
Clase Malacostraca Latreille, 1802
Infraorden Brachyura Latreille, 1802
Sección Heterotremata Guinot, 1977
Superfamilia Carpilioidea Ortmann, 1893
Familia Zanthopsidae Vía, 1959
Género Harpactoxanthopsis Vía, 1959
Harpactoxanthopsis cf. quadrilobatus (Desmarest, 1817)
(Fig. 6e-f)
Material: Un caparazón con el área ventral algo incompleta, proveniente de Urbasa 2 (sierra de Urbasa, Navarra), zona de límite entre las unidades cartográficas 268 (Formación Urbasa-Andia) y 267, Mapa Geológico de Navarra 1:25:000, Bartoniense–Priaboniense (?).
Dimensiones: L = 56,1; A = 66,9; L/A= 0.84
Descripción: Se trata del caparazón de una hembra, con el área dorsal sin cutícula y sutilmente erosionada. El caparazón es convexo, con 16 mm de grosor entre el área dorsal (parte correspondiente a la zona gástrica) y el abdomen. El margen anterior tiene un contorno semicircular (incluyendo el margen frontal y los márgenes anterolaterales). El margen anterolateral derecho presenta al menos tres bases de espinas preservadas. Conserva asimismo las bases de la espina orbital exterior izquierda y de la espina superior del margen anterolateral izquierdo. El margen posterior está parcialmente preservado. El telson es semicircular, amplio (14 mm de ancho, 10, 5 mm de largo), con una protuberancia axial bordeada por una suave depresión. El esternito 4 exhibe una protuberancia aguda alargada junto a la coxa del pereiópodo 1, el cual también presenta una protuberancia aguda, pero puntual. Maxilípedos del par 3 con isquion amplio, el cual presenta una proyección aguda, que es la base de un comparativamente estrecho exópodo.
Observaciones:Aun a pesar de su deterioro dorsal, este caparazón presenta suficientes características ─contorno, dimensiones, convexidad y espinas anterolaterales y frontales─ como para asignarlo al género Harpactoxanthopsis. Teniendo en cuenta que el grado de erosión del área dorsal no es grande y que no se observan surcos preservados, la ornamentación del mismo tuvo que estar muy poco marcada, característica asimismo típica del mencionado género. La morfología del telson, del esternito 4 y de los maxilípedos del par 3, así como las características arriba expuestas lo hacen comparable a los ejemplares de Harpactoxanthopsis quadrilobatus (Desmarest, 1817), figurados en López-Horgue & Bodego (2017; figs. 12 C-H) del Luteciense basal de la Sierra de Andia (Navarra). El fósil de Urbasa presenta un menor grosor y abombamiento del caparazón, características de los individuos adultos (e.g. Vía, 1959). Con todo, la no preservación de su cutícula dorsal (ver López-Horgue & Bodego, 2017) impide, en nuestra opinión, una asignación específica firme.
Filo Echinodermata Bruguière, 1791 (ex Klein, 1734)
Clase Crinoidea Miller, 1821
Subclase Pentacrinoidea Jaeckel, 1918
Superorden Articulata Miller, 1821
Orden Isocrinida Sieverts-Doreck, 1953
Familia Isselicrinidae Klikushin, 1977
Isocrinidae Gislén, 1924
Subfamilia Isselicrininae Klikushin, 1977
Género Isselicrinus Rovereto, 1914 (emend. Rasmussen, 1954)
Especie tipo: Isselicrinus insculptus Rovereto, 1914, por monotipia =Pentacrinites didactylus d’Orbigny en d’Archiac, 1846
Isselicrinuscf.didactylus (d’Orbigny en d’Archiac, 1846)
(Fig. 6g-i)
1846 Pentacrinites didactylus – d’Orbigny en d’Archiac: 200, pl. 5, figs. 16–18.
Material:Veintitrés fragmentos de pedúnculos, provenientes de Tiebas (Tebas-Muru Artederreta, Navarra), parte baja de la Formación Margas de Pamplona, Bartoniense. Muchos de los ejemplares están deformados.
Descripción: Pedúnculo robusto, de sección débilmente pentalobulada a subpentagonal. Debido al estado fragmentario no es posible conocer la longitud de los internodos. Los fragmentos más largos conservados tienen hasta 16 artejos o columnares. Los nodales son de igual tamaño que los internodales, con tres facetas o bases cirrales, pequeñas, de contorno subtriangular abombado, apuntando oblicuamente hacia abajo. Las articulaciones internodales son simplexiales, dando perfiles laterales crenulados. Las crenulaciones (crenularium) conforman facetas articulares con estructuras elípticas petaloides, con anchas areolas en forma de lágrima. El canal axial es pequeño, circular y centrado. Las placas columnares están ornamentadas lateralmente con pequeños tubérculos aislados y, sobre todo, alineados en su zona central, paralelos a las articulaciones y más desarrollados en las zonas radiales o interpetaloideas de cada artejo.
Observaciones: Los fragmentos de tallo de estos grandes isocrínidos son comunes en algunos afloramientos de la parte inferior de la Formación Margas de Pamplona. Su presencia en las margas eocenas de la localidad de Tiebas ya fue indicada por Ruiz de Gaona (1947a) bajo el nombre de Pentacrinus sp., el mismo que figura en la etiqueta original de la caja que conserva los ejemplares aquí descritos. Años más tarde, Ruiz de Gaona & Colom (1950: 329) indican que: “Las formaciones profundas de Tiebas ─que ubican en la que denominan serie inferior de las margas azules de Pamplona─ contienen numerosos trozos de tallos de Crinoideos del grupo de los Pentacrínidos. Carez los menciona entre Pamplona (ciudad) y la estación del ferrocarril del Norte. Sus aguas profundas no fueron aptas para el desarrollo de una fauna litoral”. La presencia de Isselicrinus en las Margas de Pamplona ha sido constatada por Astibia et al. (2005, 2016), quienes mencionan y figuran fósiles de este taxón provenientes de la localidad de Uztarrotz (Eguesibar). Un ejemplar donado por un aficionado a uno de los autores (H.A.) también permite constatar su presencia en los afloramientos margosos al pie del monte Ezkaba, al norte de Pamplona.
Los lirios de mar isocrínidos son el grupo principal de equinodermos fijos post-paleozoicos. Isselicrinus es un género extinto de la familia Isselicrinidae, que existió desde el Cretácico Superior hasta el Mioceno (Klikushin, 1982) y tuvo una amplia distribución geográfica, en los dos hemisferios. Su pedúnculo se caracteriza por tener de 1 a 3 cirros en el borde inferior de los columnares nodales, a diferencia de otros isocrínidos donde los cirros pueden ser 5 por nodal (Fujiwara et al., 2005; Kirilova & Georgiev, 2018). El número de internodales es generalmente superior a 20 —no menos de 30, según Klikushin (1982)—, a diferencia, por ejemplo, del género eoceno Cainocrinus, con nodales siempre con 5 cirros, y un número de internodales inferior a 15 (Roux et al., 2006). Sus largos internodos y menos numerosos y más espaciados cirros sugieren un modo de vida diferente al de otros Isocrinida. Isselicrinus se considera un crinoide adaptado a vivir en medios fangosos de baja energía. (Kirilova & Georgiev, 2018).
La ausencia de elementos del cáliz entre el material disponible para este trabajo dificulta la asignación taxonómica de los fósiles aquí estudiados. La morfología general de los columnares, con tres facetas o cicatrices cirrales en los nodales, y los largos internodos (hasta 16 internodales en uno de los ejemplares), son coherentes, en principio, con su asignación taxonómica a Isselicrinus. Siguiendo a Roux et al. (2006), quienes estudian los fósiles de crinoideos pedunculados de las margas del Eoceno de Miretrain (cuenca de Aquitania, área norpirenaica), los fósiles de Tiebas corresponden a morfotipos balanocrínidos. También los provenientes de Uztarrotz (Astibia et al., 2016). Sus contornos articulares internodales son como los de los (eco)fenotipos que los mencionados autores asignan a Isselicrinus sp. 4 e Isselicrinus sp. 5 —quizás más cercana a Isselicrinus sp. 4—, afines a las especies I. didactylus (d’Orbigny en d’Archiac, 1846) e I. subbasaltiformis (Miller 1821) (ver Roux et al., 2006, fig. 3). Pero debe tenerse en cuenta que, según diversos autores, el contorno del pedúnculo de Isselicrinus cambia de forma (pedúnculo xenomórfico), de estrellado en la parte proximal, pasando a pentagonal en la parte media del tallo y a casi circular en la parte distal (Kirilova & Georgiev, 2018 y otros trabajos anteriores). Según Rasmussen (1972), I. didactylus tendría columnares de mayor diámetro que I. subbasaltiformis y, además, provistos de granulación exterior, ausente en la segunda especie. Estas mismas características se observan en los fósiles de Tiebas, —con diámetros pedunculares de entre 7,3 y 8,1 mm, frente a los columnares de varias localidades del Eoceno de Dinamarca e Inglaterra (Rasmussen, 1972, fig. 4) que miden entre 3 y 7 mm — y marcada granulación exterior. Los ejemplares de Tiebas parecen similares a los fósiles originales de la especie I. didactylus, provenientes de Biarritz (Costa Vasca, cuenca de Aquitania), descritos y figurados en d’Archiac (1846, pl. 5, figs. 16-18). De Bouillé (1876) cita esta especie en los yacimientos de Abattoir (“gisement de Pentacrinites”, de donde proviene el material tipo) y en el de Lady Bruce ─llamado Villa Marbella, en Boussac (1911) ─ en la Formación Côte des Basques (Mathelin & Sztràkos, 1993) de Biarritz.
Familia Isocrinidae Gislén, 1924
Subfamilia Metacrininae Klikushin, 1977
Género Metacrinus Carpenter, 1884
Especie tipo: Metacrinus wyvillei Carpenter, 1884
Metacrinus sp.
(Fig. 6j-l)
Material: Doce artejos o placas columnares aisladas y 69 fragmentos de pedúnculos (2–10 columnares), provenientes de Ardanatz 2 (Eguesibar, Navarra), parte alta de la Formación Margas de Pamplona o parte baja de la Formación Margas de Ilundain, Bartoniense
Descripción: Pedúnculo de sección pentaestrellada a pentalobulada. Los internodos son cortos, intercalándose de 7 a 9 artejos o columnares internodales. Los columnares nodales son mayores que los internodales, con cinco facetas o bases cirrales bien marcadas y grandes, de contorno elíptico. Las articulaciones nodal-infranodales son criptosimplexiales; las demás articulaciones son de tipo simplexial, dando perfiles laterales crenulados. Las crenulaciones (crenularium) conforman facetas articulares con estructuras elípticas petaloides.
Observaciones: Ruiz de Gaona guardaba este material como Pentacrinus sp. La ausencia de elementos del cáliz entre los fósiles disponibles de Ardanatz dificulta la asignación taxonómica de los mismos. Según Klikushin (1982), en Metacrinus el tallo es de sección pentangular o pentalobulada. Los nodales son más grandes que los internodales y poseen cinco facetas cirrales profundas dirigidas hacia afuera. El número de internodales es de 7 a 12. Las crenulaciones marginales son grandes. Los pétalos son pequeños y tienen forma lanceolada. En principio, la morfología peduncular de los fósiles navarros se ajusta a estas mismas características. Metacrinus Carpenter, 1884 es un género cercano morfológicamente a los Isocrinidae de los géneros Isocrinus von Meyer en Agassiz, 1836 y Chladocrinus Agassiz, 1836. Según Hess & Messing (2011) Chladocrinus sería un subgénero de Isocrinus. Las facetas articulares son en los tres casos bastante similares. Sin embargo, siguiendo a Whittle et al. (2018), mientras que tanto en Isocrinus como en Chladocrinus el tamaño de los columnares nodales e internodales es similar, en Metacrinus, como ya se ha indicado y también es el caso del material aquí descrito, los artejos nodales son mayores, más anchos y altos, que los internodales. Además, Chladocrinus tiene internodos más largos, compuestos por más de 20 columnares. La presencia de este taxón no había sido citada hasta el momento en el Eoceno de Navarra.
Clase Echinoidea Leske, 1778
Orden Camarodonta Jackson, 1912
Familia Triplacidiidae (nombre provisional)
Género Triplacidia Bittner, 1891
Especie tipo: Micropsis biarritzensis Cotteau, 1863
Triplacidia sp.
(Fig. 7a-b)
|
 Fig. 7.— Fósiles del Eoceno [Bartoniense–Priaboniense (?)] de Navarra de la Colección Ruiz de Gaona. Triplacidia sp., CRG.52 (sigla original: 1387), vista oral (a), Urbasa 2.; CRG. 53, vista oral (b), Urbasa 2. Woellsteinia cf. kozlovi Adnet, 2006, CRG.54, diente anterior inferior (?), vistas labial (c) y lingual (d), Ardanatz (1 o 2); CRG.55, diente anterolateral superior, vistas labial (e), lingual (f) y distal (g), Sarriguren; CRG.56, diente lateral superior, vistas labial (h), lingual (i) y distal (j), Sarriguren. Osículos de equinodermos asteroideos, CRG.57, osículo o placa abactinal (k), Ardanatz 3; CRG.58, osículo marginal, vistas abactinal (l) y lateral (m), Ardanatz 3. Escalas a-b, 10 mm; c-m, 5 mm. Fig. 7.— Fósiles del Eoceno [Bartoniense–Priaboniense (?)] de Navarra de la Colección Ruiz de Gaona. Triplacidia sp., CRG.52 (sigla original: 1387), vista oral (a), Urbasa 2.; CRG. 53, vista oral (b), Urbasa 2. Woellsteinia cf. kozlovi Adnet, 2006, CRG.54, diente anterior inferior (?), vistas labial (c) y lingual (d), Ardanatz (1 o 2); CRG.55, diente anterolateral superior, vistas labial (e), lingual (f) y distal (g), Sarriguren; CRG.56, diente lateral superior, vistas labial (h), lingual (i) y distal (j), Sarriguren. Osículos de equinodermos asteroideos, CRG.57, osículo o placa abactinal (k), Ardanatz 3; CRG.58, osículo marginal, vistas abactinal (l) y lateral (m), Ardanatz 3. Escalas a-b, 10 mm; c-m, 5 mm.
|
|
Material: Dos ejemplares (uno de ellos con el número de colección 1387), aplastados, incompletos en la superficie aboral, de Urbasa 2 (sierra de Urbasa, Navarra), zona de límite entre las unidades cartográficas 268 (Formación Urbasa-Andia) y 267, Mapa Geológico de Navarra 1:25:000, Bartoniense–Priaboniense (?).
Dimensiones: CRG 52, D = 110,7 / CRG 53, D = 81,3
Descripción: Equinoideo regular con caparazón de tamaño grande, de perfil lateral hemiesférico achatado y abombado. El contorno del ámbito es circular, tendente a ligeramente pentagonal. Las placas ambulacrales son trigeminadas, con uno o dos tubérculos primarios y pares de poros que se disponen más o menos serialmente, paralelos, al peristoma. Las placas interambulacrales son anchas, con hileras horizontales de hasta cinco tubérculos primarios subiguales. Los tubérculos son imperforados y crenulados. La zona interradial presenta filas de pequeños gránulos. El deterioro de la superficie aboral impide precisar sobre la morfología del disco apical.
Observaciones: Los dos ejemplares de Urbasa 2 están etiquetados en la CRG como Triplacidia van den Heckei Agass. [=Triplacidia vandenheckei (Agassiz en Agassiz & Desor, 1846]. Triplacidia vandenheckei fue inicialente asignada al género Salmacis Agassiz, 1841, a partir de ejemplares provenientes de las margas del Eoceno (Priaboniense; Bodelle et al., 1968) de La Fontaine-du-Jarrier (Alpes-Maritimes, Provenza). Carez (1881, Pl. 7, fig. 5) figura un ejemplar de este género que clasifica como “Salmacis Van den Eckei”, proveniente de la localidad catalana de Sant Fruitós de Bages, que se conserva en el MNHN de París (MNHN.F.J00517). El género Triplacidia fue definido por Bittner (1891), quien asigna al mismo las especies T. biarritzensis (Cotteau, 1863), del Eoceno de Biarritz (cuenca de Aquitania, área norpirenaica), T. fraasi (Loriol, 1880) del Eoceno de Egipto, T. stachei (Bittner, 1880) del Eoceno de Skradin en Dalmacia, T. veronensis (Bittner, 1883) del Eoceno de Verona y T. lorioli (Cotteau, 1883) del Eoceno de Hyderabad (India), especies todas ellas asignadas anteriormente al género Micropsis Cotteau en Leymerie & Cotteau, 1856. Cotteau (1863) indica la riqueza en especies de equínidos del “nummulítico” de los acantilados de Biarritz, sumando 43 las especies estudiadas por dicho autor, la mayor parte de ellas —incluida T. biarritzensis— en afloramientos de la que él denomina “zone à Ostrea rarilamella”, dentro de las margas y calizas con Rotularia spirulaea (“Marnes et calcaires à Serpula spirulaea”). Estos afloramientos, como el de Villa Marbella y otros ubicados en la parte alta de la Formación Marnes de la Côte des Basques (Boussac, 1911; Mathelin & Sztràkos, 1993), contienen unas faunas de edad Bartoniense-Priaboniense comparables —aunque más ricas— a las de la cuenca de Pamplona (sinclinal Aranguren-Itzaga) (Astibia et al., 2016, 2018) y, a tenor de los datos aquí presentados, también a los afloramientos de Urbasa-Andia aquí mencionados. El estado incompleto de los ejemplares disponibles, junto con la presencia de sedimento fuertemente cementado adherido a los mismos dificulta su asignación específica. Triplacidia se distribuye en el Eoceno del sur de Europa, norte de África, la península arábiga y la India (The Echinoid Directory, NHM, http://www.nhm.ac.uk/our-science/data/echinoid-directory).
Este trabajo constituye la primera cita del equinoideo Triplacidia en la zona de estudio.
Filo Chordata Haeckel, 1874
Clase Chondrichthyes Huxley, 1880
Subclase Elasmobranchii Bonaparte, 1838
Cohorte Euselachii Hay, 1902
Subcohorte Neoselachii Compagno, 1977
Superorden Galeomorphii Compagno, 1973
Orden Lamniformes Berg, 1958
Familia Mitsukurinidae Jordan, 1898
Género Woellsteinia Reinecke, Stapf & Raisch, 2001
Especie tipo: Woellsteinia oligocaena Reinecke, Stapf & Raisch, 2001
Woellsteinia cf. kozlovi Adnet, 2006
(Fig. 7c-j)
2006 Woellsteinia kozlovi – Adnet: 69–71, pl. 23, figs. 1–7.
2014 Woellsteinia kozlovi – Carlsen & Cuny: 45–46, fig. 3M–R.
2016 Woellsteinia kozlovi – Astibia et al.: fig. 5z.
Material: Cuatro especímenes provenientes de Sarriguren (Navarra), parte alta (?) de la Formación Margas de Pamplona, Bartoniense. Dos especímenes provenientes de Ardanatz (Navarra). En la etiqueta se indica “izquierda, carretera a Ardanaz”. Puede corresponder a las localidades aquí denominadas como Ardanatz 1 o Ardanatz 2, o bien un lugar cercano a estas, parte alta de la Formación Margas de Pamplona o parte baja de la Formación Margas de Ilundain, Bartoniense.
Descripción: Dientes de tamaño medio. Uno de los dientes (CRG.54) (Fig. 7c-d), que corresponde a una posición anterior, conserva la corona y parte de la raíz, alcanzando una longitud apicobasal de 18 mm de altura. La corona es triangular, alta, no muy ancha en la base, y prácticamente recta. La cara labial es prácticamente plana, con una ligera concavidad en la base. La superficie de la corona lingual es muy convexa. A pesar de que la vitrodentina está muy alterada por meteorización, aún se observan unos finos pliegues axiales en el tercio proximal de la cara lingual, que desaparecen apicalmente. Los bordes cortantes son lisos y alcanzan hasta la base de la corona. Una banda lingual separa la corona de la raíz. Esta apenas conserva la parte situada bajo la base de la corona. Aún así se observa que la raíz, en origen bilobulada, es baja con una suave protuberancia lingual. El contorno basal de la raíz es regularmente curvo en vistas labial y lingual.
Otro diente también de una posición anterior, aunque de la mandíbula superior, es el ejemplar CRG.55 (Fig. 7e-g). Mide 19 mm de altura, e igualmente ha perdido la mayor parte de la raíz. La corona es triangular, alta y labiolingualmente aplanada. Las caras labial y lingual son, respectivamente, prácticamente plana y ligeramente convexa. La última está ornamentada con unos finos pliegues axiales, alguno de los cuales ocupa dos tercios de la corona. Los bordes cortantes son lisos, y se interrumpen abruptamente en la base de la corona por rotura reciente. El borde mesial es regularmente convexo, mientras que el borde distal presenta forma sigmoidea. En vista lateral, la corona es recurvada labialmente en la parte apical. Este diente presenta una banda lingual en el límite corona-raíz, con forma angular en su parte media. La raíz es baja con una suave protuberancia lingual; una fractura reciente impide confirmar la presencia de surco nutricio.
El ejemplar CRG. 56 (Fig. 7h-j), con una longitud apicobasal cercana a los 9 mm, corresponde a un diente lateral superior. Presenta una corona triangular, que se inclina distalmente, basalmente ancha y más baja que la de los dientes anteriores y un talón que se extiende hasta el límite lateral del lóbulo de la raíz. En vista mesial o distal, la corona es recta. El borde cortante distal, que es liso, continúa por el talón distal, dibujando un perfil cóncavo en vistas lingual/labial. La cara labial es plana, y tiene una depresión triangular media, poco profunda, con algunos pliegues cortos en la base de la corona. La cara labial es ligeramente convexa. El límite corona/raíz forma un amplio ángulo obtuso. La raíz, que es baja, presenta dos lóbulos separados por un ángulo obtuso. El margen distal del lóbulo distal es redondeado (falta parte del lóbulo mesial). La protuberancia lingual es amplia, existiendo un corto surco nutricio. El contorno basal de la raíz es regularmente curvo en vistas labial y lingual.
Observaciones: A partir de material fragmentario, y en su mayor parte mal conservado, Ruiz de Gaona identificó los dientes de este mitsukurinido de Sarriguren (Navarra) como Scapanorhynchus, basándose en el tamaño, forma general y la existencia de ornamentación en la cara lingual de la corona. Sin embargo, este género se diferencia por presentar unos pliegues axiales más desarrollados, la existencia de cuspidillas en dientes laterales, y en general, un diseño radicular muy diferente. Además, el registro de Scapanorhynchus se limita al Cretácico (Aptiense–Maastrichtiense), con una amplia distribución mundial (Cappetta, 2012).
Los dientes de Woellsteinia, que al carecer de cuspidillas se separan fácilmente de los de Odontaspididae, presentan una depresión triangular basal y hombros a cada lado de la corona, ornamentados con pequeños pliegues en la cara labial y finos pliegues axiales en la cara lingual (Reinecke et al., 2001). Cappetta (2012) incluye cuatro especies pertenecientes al género Woellsteinia, con un rango estratigráfico que va desde el Ypresiense superior/Luteciense al Rupeliense. Woellsteinia kozlovi Adnet, 2006 es la especie más antigua conocida del género (Ypresiense superior–Luteciense) proveniente del suroeste de Francia (Adnet, 2006) y Dinamarca (Carlsen & Cuny, 2014). La corona, con pliegues muy finos en la cara labial, presenta cúspides flanqueadas con talones casi horizontales que llegan casi al margen de la raíz en los dientes laterales. La ausencia de ornamentación en la vitrodentina de la cara lingual entra dentro de la diagnosis de la especie (Adnet, 2006).
Otras dos especies eocenas han sido registradas en la cuenca de Karakia, península de Mangyshlak, suroeste Kazajistán (Asia central): Woellsteinia hermani (Kozlov en Zhelezko & Kozlov, 1999) en rocas del Bartoniense inferior (Formación Shorym) y Woellsteinia palatasi (Kozlov, 2001) en el Bartoniense superior (parte alta de la Formación Shorym). Los dientes de ambas presentan un aspecto muy similar a los de W. kozlovi, pero son más robustos. Además, los ejemplares de W. hermani son fácilmente identificables porque carecen de ornamentación en la cara lingual de la corona (Carlsen & Cuny, 2014).
La especie más moderna del género es W. oligocaena Reinecke, Stapf & Raisch, 2001 hallada en rocas de la Formación Alzey (Rupeliense, Oligoceno inferior) de la cuenca de Mainz (Alemania). Los dientes de esta especie son, en su aspecto general, más robustos que los de W. kozlovi, los pliegues axiales en la cara lingual de la corona aparecen muy marcados—al menos en los dientes de las series anteriores— y el extremo de los lóbulos de la raíz es generalmente redondeados. Reinecke et al. (2001: 19) sugieren que W. oligocaena está también presente en la Formación Shorym del suroeste de Kazajistán.
En un contexto paleogeográfico, el género Woellsteinia está principalmente restringido a los márgenes europeos del Tetis occidental y el Peritetis oriental (mar de Turan, Asia central) durante el Eoceno. También está presente en el Paleoceno (Daniense–Selandiense) de Nueva Zelanda (Mannering & Hiller, 2008), aunque es rara con solo tres dientes recolectados. Hay que tener en cuenta que estos selacios oceánicos pudieron recorrer grandes distancias, alcanzando una distribución antitropical, una característica que se produce también en otros tiburones lamniformes.
Algunos dientes de mitsukurinidos del Bartoniense superior (Formación Margas de Ilundain, sección de Arraitza, oeste de la cuenca de Pamplona) han sido referidos a Woellsteinia kozlovi en Astibia et al. (2016). Pero dado el estado de conservación y el limitado número de especímenes disponibles en la colección de Ruiz de Gaona, el material aquí descrito se identifica como Woellsteinia cf. kozlovi.
Otros taxones no descritosTOP
La CRG contiene fósiles de algunos taxones más. Se trata de ejemplares de anélidos tubícolas (Annelida Lamarck, 1809) y briozoos (Bryozoa Ehrenberg, 1831) encostrantes de Anderatz y Urbasa, mencionados en el apartado de Procesos tafonómicos (Figs. 4c, l, 5f), así como fragmentos de conchas de moluscos escafópodos (Scaphopoda indet., Fig. 8) y osículos de equinodermos asteroideos (Asteroidea indet.) de Ardanatz 3 y Anderatz (Figs. 7k-m, 8). En total contabilizarían al menos cuatro taxones más y su estudio se pospone de momento.
|
 Fig. 8.— Unidades litoestratigráficas del Eoceno [Bartoniense–Priaboniense (?)] de la cuenca de Pamplona y de la zona de Estella-Urbasa (Navarra, área surpirenaica occidental) y distribución de los taxones fósiles de la Colección Ruiz de Gaona descritos en este trabajo. Las localidades se han ordenado en función de las unidades en las que se incluyen, pero su distribución en esta figura no debe interpretarse en términos estrictos de sucesión estratigráfica. Salvo los fósiles de esponjas y probablemente también los de corales, los fósiles de Ardanatz 3 provienen de la parte media y alta de las Areniscas de Ardanatz, en niveles estratigráficamente más altos que las localidades de Ardanatz 1 y 2, Ilundain y Oritz. Las diferencias entre Urbasa 1 y 5 y Urbasa 3 y 4, respectivamente, pueden ser mínimas o inversas a como se indican en la figura. Fig. 8.— Unidades litoestratigráficas del Eoceno [Bartoniense–Priaboniense (?)] de la cuenca de Pamplona y de la zona de Estella-Urbasa (Navarra, área surpirenaica occidental) y distribución de los taxones fósiles de la Colección Ruiz de Gaona descritos en este trabajo. Las localidades se han ordenado en función de las unidades en las que se incluyen, pero su distribución en esta figura no debe interpretarse en términos estrictos de sucesión estratigráfica. Salvo los fósiles de esponjas y probablemente también los de corales, los fósiles de Ardanatz 3 provienen de la parte media y alta de las Areniscas de Ardanatz, en niveles estratigráficamente más altos que las localidades de Ardanatz 1 y 2, Ilundain y Oritz. Las diferencias entre Urbasa 1 y 5 y Urbasa 3 y 4, respectivamente, pueden ser mínimas o inversas a como se indican en la figura.
|
|
Procesos tafonómicosTOP
El hecho de que los fósiles de este estudio pertenezcan a una colección y, por tanto, estén fuera de contexto geológico, impide llevar a cabo un análisis tafonómico detallado de los mismos y sus yacimientos. El afloramiento de Zabaldika corresponde a materiales turbidíticos marinos profundos (Payros et al., 2006, 2007) y la presencia en el mismo de un fósil de Gryphaeidae (Pycnodonte rarilamella, Fig. 5a-b) es muy probablemente consecuencia de su transporte talud abajo desde áreas someras a favor de flujos gravitacionales. Los fósiles de crinoideos de Tiebas (Fig. 6h-i) son fragmentarios y están frecuentemente deformados, pero por el contexto litológico en el que se incluyen —niveles margosos de ambiente bastante profundo— seguramente corresponden a organismos démicos, descartándose procesos importantes de remoción tafonómica. Los ejemplares de Ardanatz 3 provienen muy probablemente de niveles estudiados por Astibia et al. (2014). El fósil de esponja CRG.11 (Figs. 2a-b) proviene casi con seguridad de la parte baja de la sección AD3 de Ardanatz-Eguesibar, en donde abundan los fósiles de esponjas. Estos presentan por lo general una fragmentación intensa, pero a menudo aparecen especímenes completos o casi completos, sin evidencias de abrasión (Astibia et al. (2014). Probablemente correspondan a organismos démicos, habitantes de ambientes relativamente profundos y a elementos tafonómicos autóctonos o parautóctonos (sensu Fernández López, 1990, 1991), depositados en un ambiente de baja energía. Por el contrario, en los niveles con alto contenido terrígeno de otros afloramientos de Ardanatz (Ardanatz 3 en este trabajo) el contenido macropaleontológico (fósiles de macroforaminíferos, corales, moluscos y otros organismos) también es localmente abundante, pero en estos casos los restos parecen haber sufrido procesos de remoción tafonómica (sensu Fernández López, 2000). Los macrofósiles corporales en Ardanatz están generalmente incompletos, con evidencia de disolución y reemplazo por calcita y celestita en esponjas silíceas y disolución, neomorfismo de aragonito-calcita y reemplazo por celestita en corales y moluscos (Astibia et al., 2005; 2014; 2018).
En los fósiles de la localidad de Anderatz (Rotularia spirulaea, Pycnodonte, Chlamys, Spondylus y otros, Figs. 4b, 4l-m, 5d-f) las estructuras y microestructuras, originariamente de composición carbonatada, se reconocen de manera incompleta al haber sufrido durante la diagénesis temprana una intensa silicificación, con remplazamiento por esferulitos de cuarcina-lutecita y micro-megacuarzo y formación de conjuntos superficiales de anillos de beekita (Elorza & Astibia, 2018).
La silicificación también afecta a los ejemplares de briozoos ciclostomados (Cyclostomatida indet.) y Rotularia spirulaea de los afloramientos de la sierra de Urbasa (Figs. 3e-l, 4a). Esta silicificación parece estar potenciada por la temprana degradación microbiana aeróbica de la materia orgánica en el interior del esqueleto, donde se crea un microambiente altamente restringido, con acidificación creciente y disolución del carbonato de calcio (Butts, 2014; Elorza & Astibia, 2018).
Bioerosión y epibiontesTOP
En tres ejemplares de Dimya pamplonensis se observan señales de depredación por gasterópodos (Oichnus isp., Fig. 5h-i). En Ardanatz los epibiontes (principalmente tubos de anélidos y colonias de briozoos encostrantes) y las señales de bioerosión (principalmente perforaciones de briozoos, Pinaceocladichnus isp.) sobre los corales, Rotularia spirulaea y las conchas de moluscos son frecuentes (Astibia et al., 2014, fig. 11j-k; Elorza & Astibia, 2017, fig. 2b, e). Las colonias de briozoos encostrantes son también frecuentes en los fósiles de Anderatz (Fig. 4l-m). En algunos fósiles de Rotularia spirulaea y de Pycnodonte brongniarti recogidos recientemente en Anderatz por uno de los autores (H.A.) se reconocen señales de bioerosión por briozoos (?Pinaceocladichnus isp.) y, sólo en P. brongniarti, también por esponjas (Entobia isp.). Sendos ejemplares de Rotularia spirulaea provenientes de los afloramientos de Urbasa 1 y 5 presentan en su superficie fósiles de colonias multiseriales de briozoos encostrantes (Fig. 4c). Sobre algunos ejemplares de las colonias de briozoos multilamelares de Urbasa se observan perforaciones que podrían atribuirse a anélidos poliquetos (?Caulostrepsis) (Fig. 3i).
Biodiversidad y sesgos tafonómicos en las faunas del Eoceno de NavarraTOP
La muestra estudiada en este trabajo es poco diversa, pues tan solo incluye unos 28 taxones, 24 de los cuales se han descrito aquí con cierto detalle, cuya distribución se indica en la figura 8. Además, se han incluido dos de los al menos cuatro taxones no descritos (ver apartado de Otros taxones no descritos). Astibia et al. (2014, 2016, 2018) han subrayado la menor diversidad que parecen mostrar las asociaciones fósiles del Eoceno de la cuenca de Pamplona y alrededores (sinclinal de Aranguren-Itzaga) en comparación con las contemporáneas de otras zonas surpirenaicas. Villalta Comella (1956) describe 81 especies de gasterópodos en el Eoceno del Valle de Basa, en la cuenca de Jaca (Aragón, Formación Margas de Pamplona; geología en Brzobohatý & Nolf, 2011). También con respecto a las asociaciones del Eoceno de Biarritz, en el área norpirenaica (cuenca de Aquitania), donde Boussac (1911) describe y/o cita 30 especies de moluscos en el afloramiento de Villa Marbella y 93 especies en niveles suprayacentes de la Côte des Basques (Formación Marnes de la Côte des Basques, Bartoniense–Priaboniense; geología en Mathelin & Sztràkos, 1993). A su vez, la biodiversidad de estas asociaciones parece mucho menor si se las compara con las ricas faunas fósiles del Bartoniense de la cuenca de París (1000 especies de gasterópodos y 400 de bivalvos; en Merle, 2008), pero no tanto con las del Priaboniense (70 especies de gasterópodos y 50 de bivalvos; en Merle, 2008), que sufren en la cuenca parisina una caída de biodiversidad muy fuerte que se recupera en la fachada atlántica durante el Estampiense.
La menor riqueza de las asociaciones de invertebrados del Eoceno navarro puede deberse a diferencias paleoecológicas y/o sesgos tafonómicos y de muestreo. En relación con el último de estos factores, en los trabajos arriba mencionados se indica la naturaleza todavía preliminar de los estudios realizados en Navarra y lo limitado del muestreo realizado, ceñido básicamente a la recogida de fósiles sueltos sobre el regolito margoso, hecho que, por ejemplo, no ha permitido analizar el componente de micromoluscos. Es muy posible que este factor también haya influido en la constitución de la muestra obtenida por Ruiz de Gaona, cuya trayectoria investigadora estuvo centrada en los fósiles de foraminíferos.
Por otro lado, también es muy probable la existencia de un importante sesgo tafonómico a favor de los esqueletos calcíticos en detrimento —por disolución— de los fósiles de organismos con esqueletos aragoníticos. La disolución temprana del aragonito en ambientes de plataforma y cuenca constituye un importante factor en el sesgo del registro fósil (Wright et al., 2003; Cherns et al., 2011). Wright et al. (2003) constatan este hecho refiriéndose a faunas marinas del Jurásico Inferior de Gales y sugieren que los procesos de disolución de conchas aragoníticas ocurrieron principalmente en entornos de baja energía, donde los altos contenidos orgánicos habrían favorecido la descomposición microbiana y la acidez del medio. Según Cherns et al. (2011) la disolución de las conchas de moluscos en la zona tafonómicamente activa del fondo marino (en inglés TAZ), es debida a una subsaturación causada por la degradación microbiana de la materia orgánica y otras reacciones relacionadas. Sanders et al. (2015) estudian el efecto de la litificación (diagénesis) en las capas del Eoceno medio (Luteciense medio) del “Falunière” de Grignon (Yvelines), en la cuenca de París, llegando a la conclusión de que este proceso provoca un importante problema (sesgo) a la hora de evaluar la paleobiodiversidad registrada. En las facies litificadas estos autores estiman una pérdida de alrededor del 80% de la riqueza específica de los moluscos, porcentaje que llega al 100% en las especies menores de 2 mm, en comparación con las capas no litificadas de la misma localidad fosilífera.
Las extensas formaciones margosas del Bartoniense–Priaboniense marino de Navarra resultan en general muy pobres en macrofósiles y cuando —solo localmente— estos son numerosos hay un claro predominio de fósiles calcíticos. Dentro de los moluscos, los bivalvos más abundantes en los afloramientos estudiados pertenecen a la infraclase Pteriomorphia (familias Gryphaeidae, Pectinidae, Spondylidae y Dimyidae, entre otras), que son mayoritariamente de hábitos de vida epibentónico y producen en sus conchas capas calcíticas (Esteban, 2006). Otros bivalvos abundantes pertenecen a las especies del género Chama (Astibia et al., 2018 y en este trabajo). Según Esteban (2006) algunas especies de este género, —excepción entre los Heteroconchia, predominantemente endobentónicos y de concha aragonítica—, son epibentónicas y tienen una capa calcítica externa. En aquellos enclaves donde hay fósiles de gasterópodos es muy posible que sus conchas, originariamente aragoníticas, estén neomorfizadas a calcita en la mayoría de los casos. Astibia et al. (2018) indican que las conchas de Ptychocerithium baylei, especie frecuente en algunos niveles de las secciones de Ardanatz y Aranguren, están parcialmente disueltas y neomorfizadas a calcita y sus cavidades cementadas por celestita. La disolución parcial de las conchas es patente en la mayoría de los fósiles de otras especies de gasterópodos de estos afloramientos. El neomorfismo aragonito-calcita afecta asimismo a la capa interna de estructura prismática, originalmente aragonítica, de las conchas del bivalvo Spondylus cisalpinus provenientes de las secciones de Aranguren e Itzagaondoa (Astibia et al., 2018).
En cuanto a otros invertebrados, los ejemplares del coral Placosmiliopsis bilobatus de la sección AD3 de Ardanatz (Eguesibar) muestran septos originalmente aragoníticos completamente neomorfizados por grandes cristales de calcita (Astibia et al., 2014). Los tubos del anélido Rotularia spirulaea, muy abundantes en algunos de los niveles bartonienses de la zona, son de composición calcítica, sin evidencias de la presencia de aragonito, y están reemplazados por sílice en el área de Anderatz-sierra de Urbasa (Fig. 4a-b; Elorza & Astibia, 2017, 2018). Los esqueletos coloniales de briozoos estenolemados también son de calcita (Taylor et al., 2015), pero en los fósiles de Cyclostomatida indet. del Bartoniense de la sierra de Urbasa estos están igualmente reemplazados por silice. Si bien el contenido macropaleontológico del Eoceno medio y superior marino de Navarra es escaso, no lo es el micropaleontológico, rico en foraminíferos de pared aglutinante y, sobre todo, Rotaliina, de pared calcítica hialina, además de ostrácodos, serpúlidos, espinas de equínidos y girogonitos de carofitas, entre otros elementos (Astibia et al., 2014, 2016).
Así pues, tal y como se ha indicado, la escasa diversidad de las asociaciones fósiles estudiadas en este y en anteriores trabajos, puede ser más debida a un efecto tafonómico de disolución que a causas de naturaleza ecológica.
Asociaciones fósiles, correlación y paleoambientesTOP
La evolución tectonosedimentaria y las asociaciones fósiles de la zona de estudio indican una progresiva somerización del medio durante el Eoceno, desde un mar profundo a condiciones intertidales, a una con el progresivo relleno de la cuenca de antepaís surpirenaica (Payros, 1997; Pujalte et al., 2002; Payros et al., 2010; Astibia et al., 2016, 2017).
Los materiales de Zabaldika corresponden a la transición entre los depósitos calciturbidíticos distales de la Formación Anotz, acumulados al pie de un talud carbonatado, y los depósitos turbidíticos siliciclásticos distales de la Formación Erro (Grupo Hecho), acumulados axialmente en el fondo de la cuenca surpirenaica (Payros et al., 1999, 2006). Según Payros et al. (2006, 2007, 2011) las intercalaciones margosas de la Formación Anotz contienen una rica asociación de foraminíferos planctónicos, que superan el 60% de la asociación total de foraminíferos (planctónicos y bentónicos), y que incluye tanto formas esféricas de pared delgada como formas carenadas de pared gruesa. Según estas características, dichos autores estimaron una paleoprofundidad de unos 500 m.
Los niveles margosos de la localidad de Tiebas, en la parte baja de la Formación Margas de Pamplona, corresponden todavía a un medio relativamente profundo. Datos micropaleontológicos de la localidad estratigráficamente equivalente de Uztarrotz (Eguesibar), revelan una considerable abundancia de fósiles de foraminíferos, con moderado dominio de especies planctónicas, factor que, junto con la alta diversidad y composición de la asociación de especies bentónicas, sería indicativo de un ambiente de plataforma externa o parte alta del talud continental (Astibia et al., 2016). En el mismo sentido apunta la presencia en la localidad de Tiebas, y también en Uztarrotz (Astibia et al., 2016), de fósiles de Isselicrinus, crinoides adaptados a ambientes fangosos de baja energía, principalmente restringidos a entornos de aguas profundas en el Cenozoico (Fujiwara et al., 2005; Malubián & Olivero, 2005). Roux et al. (2006) proponen una zonación batimétrica del género Isselicrinus en las margas del Eoceno de Miretrain (cuenca de Aquitania, área norpirenaica), y describen cuatro (eco)fenotipos, indicativos de profundidades entre 200 y 1000 o más metros. Los hallados en Tiebas y Uztarrotz son morfotipos balonicrínidos, adaptados a flujos de corrientes laminares, con contornos de las articulaciones internodales similares a los de las especies Isselicrinus sp. 4 y Isselicrinus sp. 5 de Roux et al. (2006), afines a las especies I. didactylus e I. subbasaltiformis, respectivamente, para los que en el mencionado trabajo asignan profundidades entre 200 y 400 metros.
Subiendo en la serie, en algunos puntos de la cuenca de Pamplona —cercanos o coincidentes con afloramientos como los de Sarriguren, Ardanatz 1, Ardanatz 2 e Ilundain, mencionados en este trabajo— las Margas de Pamplona e Ilundain se suceden sin la intercalación de las areniscas de Ardanatz. En estos lugares, así como hacia el oeste de la cuenca de Pamplona, en la sección de Arraitza (Margas de Ilundain), los depósitos aún pueden corresponder a un ambiente relativamente profundo. En la sección de Arraitza los datos micropalentológicos indican profundidades de unos 200 metros (Astibia et al., 2014). Puesto que los crinoideos pedunculados actuales viven en ambientes profundos, puede deducirse que la presencia de fósiles de crinoideos isocrínidos (Metacrinus sp.) en Ardanatz 1 y Ardanatz 2 es también indicativa de un ambiente profundo. De hecho, los isocrínidos actuales viven generalmente entre 100 y 400 metros de profundidad (Whittle et al., 2018). Con todo, diversos trabajos describen fósiles de isocrínidos y otros crinoideos (bourgueticrínidos) de comunidades que habitaron en el Cretácico y Paleógeno en aguas más someras (Whittle et al., 2018; Zamora, 2018; Zamora et al., 2018).
La existencia de dientes atribuidos al mitsukurinido Woellsteinia en Sarriguren, Ardanatz (1-2) (en este trabajo) y Arraitza (Astibia et al., 2016) indicaría un ambiente típicamente pelágico marino con niveles normales de salinidad. Sin embargo, es difícil hacer una inferencia paleobatimétrica a partir de este único género, más aún cuando se trata de un taxón extinguido. Woellsteinia está presente en distintas asociaciones de selacios del Eoceno y Oligoceno de Europa (cuencas de Aquitania y del Mar del Norte) (Adnet, 2006; Schnetler & Heilmann-Clausen, 2011; Carlsen & Cuny, 2014) y Asia central, en la península de Mangyshlak (Zhelezko & Kozlov, 1999; Pelc et al., 2011). En general, el género se asocia con ambientes de plataforma externa y talud continental. Sin embargo, Woellsteinia también aparece en el Oligoceno inferior (Rupeliense) de la cuenca de Mainz (Alemania), en una unidad transgresiva junto con fósiles de foraminíferos, moluscos, ostrácodos, selacios, peces óseos y mamíferos marinos, asociados a medios litoral y pelágico, con un rango de profundidad de 7–70 m (Schindler et al., 2009).
Como ya se ha indicado (ver apartado de Material y métodos), los fósiles de Ardanatz 3 provienen muy probablemente del entorno de las secciones AD1, AD2 y AD3 de Astibia et al. (2014). En el tramo inferior de estas secciones de Ardanatz y, asimismo, en la cercana sección de Aranguren (niveles AD3.1, AD1.1, AG1.1 de Astibia et al. 2014) la litología margosa y la abundancia local de fósiles de esponjas hexactinélidas (Guettardiscyphia thiolati/Pleuroguetardia ibérica y otros taxones) y “litístidas” (Heteroscleromorpha), así como de colonias erectas y ramificadas de briozoos —también hay fósiles de crinoideos isocrínidos (Crinoidea indet., en Astibia et al., 2016)—, son indicativas de un medio relativamente profundo y de baja energía. En la parte baja de las Areniscas de Ardanatz y de las Margas de Ilundain también son localmente abundantes el braquiópodo Terebratulina tenuistriata y el bivalvo Dimya pamplonensis. Sin embargo, los tramos medio y alto de las secciones de Ardanatz, donde todos estos fósiles parecen ausentes y, por el contrario, son frecuentes los del anélido Rotularia spirulaea y los de gasterópodos como Ptychocerithium baylei y otros muchos moluscos, corresponden seguramente a un ambiente de menor profundidad. La asociación micropaleontológica en los niveles AD3.1 y AD2.1 de la sección de Ardanatz muestra un dominio de foraminíferos bentónicos, que por su abundancia y composición indicarían un ambiente marino de plataforma (Astibia et al., 2014).
En la zona occidental de este estudio la Formación de Calizas Urbasa-Andia constituye el registro de una rampa carbonatada transgresiva, marina somera, dominada por fuertes tormentas (Payros et al., 2010). Como ya se ha indicado (ver apartado de Procesos tafonómicos), los fósiles de estos niveles, generalmente fragmentarios y silicificados, provienen de restos que han sufrido remoción tafonómica. La presencia en Anderatz y también en Urbasa de las especies Rotularia spirulaea, Pycnodonte brongniarti y Spondylus cisalpinus, acerca estos depósitos occidentales desde un punto de vista paleoambiental a los tramos altos de las Areniscas de Ardanatz (R. spirulaea y S. cisalpinus ) y Margas de Ilundain (R. spirulaea, P. brongniarti y Spondylus cisalpinus) (Astibia et al., 2016, 2018), que constituyen las capas más altas del Eoceno del sinclinal Aranguren-Itzaga, de edad Bartoniense o ya Priaboniense, correspondientes a ambientes marinos de plataforma interna.
En cuanto a la correlación estratigráfica entre las dos áreas de estudio, al este y oeste de la falla de Pamplona, es probable que los depósitos más altos de la sierra de Urbasa (calcarenitas y areniscas de la unidad 271 y margas de la unidad 267, Mapa Geológico de Navarra 1:25.000, http://geologia.navarra.es/) sean tan recientes como los más altos del sinclinal Aranguren-Itzaga. Basándose en criterios litológicos, según Payros et al. (2000) podrían corresponder a la Formación Gendulain. En el Mapa Geológico de Navarra los depósitos margosos de Urbasa donde se ubican los puntos fosilíferos de Urbasa 3 y 4 se incluyen en la misma unidad litológica (267) que las Margas de Pamplona, al este de la zona de estudio. Sin embargo, ni P. brongniarti (Urbasa 3) ni R. spirulaea (Urbasa 4), están presentes en las Margas de Pamplona. Contrariamente y como se acaba de indicar, ambas especies son abundantes en la parte alta de las Margas de Ilundain, suprayacentes a las de Pamplona. No obstante, estos datos pueden interpretarse en términos ambientales, indicativos de un medio marino somero en Urbasa, y de un medio más profundo en relación con las Margas de Pamplona. De hecho, ninguno de estos dos taxones permite un gran afine bioestratigráfico. P. brongniarti es una especie cuyo rango temporal abarca desde el Paleoceno hasta el Mioceno (Abad, 2001) y R. spirulaea, al menos desde el Eoceno medio (Luteciense inferior) hasta el Eoceno superior Priaboniense (Mikuž, 2008; Elorza & Astibia, 2018). A falta de una magnetoestratigrafía y bioestratigrafía (micropaleontología) de detalle, resulta difícil precisar más.
ConclusionesTOP
Con este trabajo se ha pretendido contribuir a valorar una vez más la figura y legado científico de Ruiz de Gaona y aportar nuevos datos al conocimiento del registro fósil del Eoceno pirenaico. El estudio de los fósiles de la Colección Ruiz de Gaona ha permitido describir 24 taxones de macrofauna marina del Eoceno medio–superior de Navarra. Siete de ellos ─Cyclolitopsis patera (d’Achiardi, 1867), Funginellastraea barcelonensis (Oppenheim, 1911), Pycnodonte rarilamella (Melleville, 1843), Metacrinus sp., Triplacidia sp., además de un briozoo Cyclostomatida y un anélido Serpulidae indeterminados─ son nuevos para el área de estudio. Los cuatro primeros y Serpulidae indet. provienen de la cuenca de Pamplona (sinclinal de Aranguren-Itzaga); Triplacidia sp. y Cyclostomatida indet., de la sierra de Urbasa. Se confirma la extensión de las especies Terebratulina tenuistriata (Leymerie, 1846), Dimya pamplonensis (Carez, 1881), Venericardia hortensis (Vinassa de Regny, 1897), Isselicrinus cf. didactylus (d’Orbigny en d’Archiac, 1846) y una Heteroscleromorpha indet. a otros enclaves fosilíferos. A este respecto, la revisión de la CRG incluye nuevas localidades, en concreto Zabaldika, Tebas-Tiebas, Oritz, Ilundain, Ardanatz 1 y 2, y Sarriguren, algunas mencionadas en trabajos precedentes por Ruiz de Gaona, pero de las que no existían descripciones de macrotaxones fósiles. Los afloramientos de las tres últimas localidades han desaparecido, debido al crecimiento urbano y las transformaciones agrarias.
Si bien las asociaciones macropaleontológicas del Eoceno de la cuenca de Pamplona vienen describiéndose con algún detalle en las últimas décadas, no ha ocurrido lo mismo con las existentes al oeste de la falla de Pamplona, en la Formación Calizas de Urbasa-Andia y niveles suprayacentes, prácticamente desconocidas hasta el presente. Solo las especies Rotularia spirulaea (Lamarck, 1818) y Harpactoxanthopsis sp., provenientes de Anderatz y de la sierra de Urbasa (Urbasa 2), han sido objeto de breves descripciones en sendos trabajos recientes. La lista faunística de esta área queda provisionalmente ampliada con los taxones Cyclostomatida indet., Pycnodonte brongniarti (Bronn, 1831), Chamys cf. biarritzensis (d’Archiac, 1846), Spondylus cisalpinus Brongniart, 1823 y Triplacidia sp.
Los niveles fosilíferos más antiguos aquí mencionados son los de Zabaldika (Luteciense medio), que junto con los de Tiebas, corresponden a los ambientes marinos más profundos. Los niveles más recientes y someros son probablemente los de Urbasa 3 y 4 (probablemente Priaboniense), en la parte más occidental de la zona de estudio.
El total de taxones descritos en este y en trabajos precedentes sobre el registro fósil del Eoceno medio-superior de Navarra conforman asociaciones macrofaunísticas de baja diversidad, si se las compara con las existentes en otras regiones geológicas europeas. Más allá de seguros sesgos de muestreo y posibles condicionamientos ecológicos, se plantea la existencia de un sesgo tafonómico como principal factor responsable de la misma, consecuencia de una importante pérdida en el registro de las especies de invertebrados con esqueletos aragoníticos.
AGRADECIMIENTOSTOP
A la Dra. Esther Martín (Museo de Ciencias Naturales de Tenerife) y al Dr. Julio Aguirre (Universidad de Granada), por sus aportaciones como revisores, las cuales han permitido mejorar notablemente el manuscrito. Al Dr. Javier Elorza (Universidad del País Vasco/Euskal Herriko Unibertsitatea), por la lectura crítica de algunas partes del texto. Al Dr. Paul D. Taylor (Natural History Museum, London) por su ayuda en el estudio de los briozoos. Los autores agradecen al Dr. Jesús Sesma y a Rubén Jiménez, de los Fondos de Arqueología del Departamento de Cultura, Deporte y Juventud (Sección de Arqueología, Servicio de Patrimonio Histórico) del Gobierno de Navarra-Nafarroako Gobernua, por sus facilidades y gestiones que han posibilitado estudiar los materiales fósiles de la Colección Ruiz de Gaona. Este trabajo ha sido subvencionado en parte por los grupos de investigación PPG17/05 (Universidad del País Vasco/Euskal Herriko Unibertsitatea) e IT1418-19 (Gobierno Vasco/Eusko Jaurlaritza) y el proyecto CGL2017-85038-P (Ministerio de Economía, industria y Competitividad, Servicio de Estado de Investigación, Gobierno de España, MINECO) y el Fondo Europeo de Desarrollo Regional (ERDF/FEDER). La aportación de A.P. está financiada por el grupo de investigación IT-930-16 (Gobierno Vasco/Eusko Jaurlaritza) y el proyecto CGL2015-65404-R (MINECO/FEDER, EU).
ReferenciasTOP
| ○ |
Abad, A. (2001). Paleotaxodonta y Pteriomorphia del Eoceno del Margen Sur de la Depresión Catalana. Tesis doctoral, Universitat Autònoma de Barcelona, 803 pp.
|
| ○ |
Adnet, S. (2006). Nouvelles faunes de Sélaciens (Elasmobranchii, Neoselachii) de l’Eocène moyen des Landes (Sud-Ouest, France). Implication dans la connaissance des communautés de sélaciens d’eaux profondes. Palaeo Ichthyologica, 10: 1–128.
|
| ○ |
Agassiz L. & Desor P.J.E. (1846). Catalogue raisonné des familles, des genres, et des espèces de la classe des échinodermes. Annales des Sciences Naturelles, 3ème série, Zoologie, 6: 305–374.
|
| ○ |
Agassiz, L. (1836). Prodrome d’une Monographie des Radiaires ou Echinodermes. Mémoires de la Société des Sciences naturelles de Neuchâtel, 1: 168–199.
|
| ○ |
Agassiz, L. (1841). Monographies d’Échinodermes vivans et fossiles. Première Monographie), Neuchâtel, Switzerland, i–x. https://doi.org/10.5962/bhl.title.126954 |
| ○ |
Alameda, J.; Blas, I.; Cabra, P.; Del Valle de Lersundi, J.; García, A.; Martínez, A.; Ramírez del Pozo, J. & Solé, J. (1993). Hoja geológica num. 141–II (Pamplona). Cartografía Geológica de Navarra E. 1:25.000, Gobierno de Navarra, Departamento de Obras Públicas, Transportes y Comunicaciones, Pamplona.
|
| ○ |
Alloiteau, J. (1952). Madréporaires post-paleozoiques. In: Traité de Paléontologie (Piveteau, J., Ed.), Masson, Paris, 376–684. |
| ○ |
Álvarez-Pérez, G. (1993). Cnidaria fòssils de la Conca d’Igualada. Tesis doctoral, Universitat de Barcelona, 270 pp. |
| ○ |
Álvarez-Pérez, G. (2009). Actualització de les espècies coral.lines de l’Eocè de les conques sudpirinenques. Batalleria, 14: 5–10.
|
| ○ |
Álvarez-Pérez, G. & Busquets, P. (2012). Formas anómalas en los corales eocenos de la Cuenca de Igualada (Noreste de España). Revista Española de Paleontología, 27(1): 15–28.
|
| ○ |
Archiac, A.M. d’ (1846). Description des fossiles recueillis par M. Thorent dans les couches à Nummulines des environs de Bayonne. Mémoires de la Société géologique de France, 2ème série, 2(4): 189–217.
|
| ○ |
Archiac, A.M. d’ (1847). Extrait d’un Mémoire sur les fossiles des couches à Nummulites des environs de Bayonne et de Dax. Bulletin de la Société géologique de France, 2ème série, 4(2): 1006–1015.
|
| ○ |
Archiac, A.M. d’ (1850a). Description des fossiles du groupe nummulitique recueillis par M.S.-P. Pratt et M.J. Delbos aux environs de Bayonne et de Dax. Mémoires de la Société géologique de France, 2ème série, 3(6): 397–456.
|
| ○ |
Archiac, A.M. d’ (1850b). Formation nummulitique. Roches ignées ou pyrogènes des époques quaternaire et tertiaire. In: Histoire des progrès de la géologie de 1834–1859, Société Géologique de France, sous les auspices de M. le Ministre de l’instruction publique, 3, 624 pp.
|
| ○ |
Arlegi, M.; Ríos-Garaizar, J.; Rodríguez-Hidalgo, A.; López-Horgue, M.A. & Gómez-Olivencia, A. (2018). Koskobilo (Olazti, Nafarroa): nuevos hallazgos y revisión de las colecciones. Munibe (Antropologia-Arkeologia), 69: 21–41. https://doi.org/10.21630/maa.2018.69.07 |
| ○ |
Astibia, H.; Elorza, J.; Pisera, A.; Álvarez-Pérez, G.; Payros, A. & Ortiz, S. (2014). Sponges and corals from the Middle Eocene (Bartonian) marly formations of the Pamplona Basin (Navarre, western Pyrenees): taphonomy, taxonomy, and paleoenvironments. Facies, 60: 91–110. https://doi.org/10.1007/s10347-013-0364-2 |
| ○ |
Astibia, H.; Mazo, A.V. & Santafé, J.V. (1987). Los macromamíferos del Mioceno medio de las Yeseras de Monteagudo (Depresión del Ebro, Navarra). IV Congreso Latinoamericano de Paleontología, Santa Cruz de la Sierra, Bolivia: 415–441.
|
| ○ |
Astibia, H.; Merle, D.; Pacaud, J.M.; Elorza, J. & Payros, A. (2018). Gastropods and bivalves from the Eocene marly formations of the Pamplona Basin and surrounding areas (Navarre, western Pyrenees). Geodiversitas, 40(11): 211–257. https://doi.org/10.5252/geodiversitas2018v40a11 |
| ○ |
Astibia, H.; Murelaga, X. & Pereda Suberbiola, X. (1996). Máximo Ruiz de Gaona como prehistoriador y paleontólogo de vertebrados. In: Homenaje a Máximo Ruiz de Gaona, naturalista y paleontólogo (1917–1971) (Astibia, H.; del Valle de Lersundi, J.; Murelaga, X. & Serra-Kiel, J., Coords.), Príncipe de Viana (Suplemento de Ciencias), 14, Gobierno de Navarra, Pamplona, 65–76.
|
| ○ |
Astibia, H.; Murelaga, X.; Payros, A.; Pereda, X. & Tosquella, J. (1999). Tortugas y Sirenios fósiles en el Eoceno marino de Navarra y Cuenca de Jaca. Geogaceta, 25: 15–18.
|
| ○ |
Astibia, H.; Payros, A.; Ortiz, S.; Elorza, J.; Álvarez-Pérez, G.; Badiola, A.; Bardet, N.; Berreteaga, A.; Bitner, M.A.; Calzada, S.; Corral, J.C.; Díaz-Martínez, I.; Merle, D.; Pacaud, J.M.; Pereda-Suberbiola, X.; Pisera, A.; Rodríguez-Tovar, F.J. & Tosquella, J. (2016). Fossil associations from the middle and upper Eocene strata of the Pamplona Basin and surrounding areas (Navarre, western Pyrenees). Journal of Iberian Geology, 42: 7–28. https://doi.org/10.5209/rev_JIGE.2016.v42.n1.51601 |
| ○ |
Astibia, H.; Payros, A.; Pereda Suberbiola, X.; Elorza, J.; Berreteaga, A.; Etxebarria, N.; Badiola, A. & Tosquella, J. (2005). Sedimentology and taphonomy of sirenian remains from the Middle Eocene of the Pamplona Basin (Navarre, western Pyrenees). Facies, 50: 463–475. https://doi.org/10.1007/s10347-004-0026-5 |
| ○ |
Astibia, H.; Pereda Suberbiola, X.; Bardet, N.; Payros, A.; Berreteaga, A. & Badiola, A. (2006). Nuevos fósiles de sirenios en el Eoceno medio de la Cuenca de Pamplona (Navarra). Revista Española de Paleontología, 21: 79–91.
|
| ○ |
Astibia, H.; Pereda Suberbiola, X.; Payros, A.; Murelaga, X.; Berreteaga, A.; Baceta, J.I. & Badiola, A. (2007). Bird and mammal footprints from the Tertiary of Navarre (western Pyrenees). Ichnos, 14: 175–184. https://doi.org/10.1080/10420940601049917 |
| ○ |
Astibia, H.; Rodríguez-Tovar, F.J.; Díaz-Martínez, I.; Payros, A. & Ortiz, S. (2017). Trace fossils from the Middle and Upper Eocene (Bartonian-Priabonian) molasse deposits of the Pamplona Basin (Navarre, western Pyrenees): palaeoenvironmental implications. Geological Journal, 52: 327–349. https://doi.org/10.1002/gj.2763 |
| ○ |
Barnolas, A.; Payros, A.; Samso, J.M.; Serra-Kiel, J. & Tosquella, J. (2004). La Cuenca surpirenaica desde el Ilerdiense medio al Priaboniense. In: Geología de España (Vera, J.A., Ed.), Instituto Geológico y Minero de España, Sociedad Geológica de España, Madrid, 313–320.
|
| ○ |
Barta-Calmus, S. (1973). Révision des collections de madréporaires provenant du nummulitique du Sud-Est de la Frace, de l’Italie et de la Yougoslavie septentrionales. Tesis doctoral, Université de Paris 6, 695 pp.
|
| ○ |
Bellardi, L. (1852). Catalogue raisonné des fossiles nummulitiques du Comté de Nice. Mémoires de la Société géologique de France, 2(4): 205–300.
|
| ○ |
Bellardi, L. & Michelotti, G. (1840). Saggio orittografico sulla classe dei Gasteropodi fossili dei terreni terziarii del Piemonte. Memorie della Reale Accademia delle Scienze di Torino, Serie 2(3): 93–174.
|
| ○ |
Ben-Eliahu, M.N. & Ten Hove, H.A. (2011). Serpulidae (Annelida: Polychaeta) from the Suez Canal-From a Lessepsian Migration Perspective (a Monograph). Zootaxa, 2848: 1–147. https://doi.org/10.11646/zootaxa.2848.1.1 |
| ○ |
Berggren, W.A.; Kent, D.V.; Swisher, C.C. III & Aubry, M.P. (1995). A revised Cenozoic geochronology and chronostratigraphy. In: Geochronology, time scales and global stratigraphic correlations (Berggren, W.A.; Kent, D.V.; Aubry, M.P. & Hardenbol, J., Eds.), SEPM, Society for Sedimentary Geology, Special Publications, 54: 129–212. https://doi.org/10.2110/pec.95.04.0129 |
| ○ |
Bitner, M.A. (2000). Lower Eocene (Middle Ilerdian) brachiopods from the Campo region, Central Pyrenees, north-eastern Spain. Revista Española de Paleontología, 15: 117–128.
|
| ○ |
Bitner, M.A. & Boukhary, M. (2012). First record of Eocene brachiopods from the United Arab Emirates, Arabian Gulf and their paleogeographical significance. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 265: 275–279. https://doi.org/10.1127/0077-7749/2012/0263 |
| ○ |
Bitner, M.A. & Dulai, A. (2008). Eocene micromorphic brachiopods from north-western Hungary. Geologica Carpathica, 59: 31–43. |
| ○ |
Bitner, M.A. & Müller, A. (2015). Brachiopods from the Silberberg Formation (Late Eocene to Early Oligocene) of Atzendorf, Central Germany. Paläontologische Zeischrift, 89: 673–688. https://doi.org/10.1007/s12542-015-0262-8 |
| ○ |
Bitner, M.A. & Müller, A. (2017). Late Eocene (Priabonian) brachiopod fauna from Dnipropetrovsk, eastern Ukraine. Bulletin of Geosciences, 92(2): 211–231. https://doi.org/10.3140/bull.geosci.1661 |
| ○ |
Bitner, M.A.; Astibia, H. & Payros, A. (2016). Middle Eocene (Bartonian) brachiopods from the Pamplona Basin, Navarre, South-Western Pyrenees. Batalleria, 23: 1–7.
|
| ○ |
Bittner, A. (1880). Beiträge zur Kenntnis alttertiärer Echinidenfaunen der Südalpen. Beiträge zur Paläontologie Oesterreich Ungarns. I, Wien.
|
| ○ |
Bittner, A. (1883). Micropsis Veronensis, ein neuer Echinide des oberitalienischen Eocaens. Sitzungsberichte der Akademie der Wissenschaften mathematisch-naturwissenschaftliche Klasse 88: 444–449.
|
| ○ |
Bittner, A. (1891). Ueber Parabrissus und einige andere alttertiäre Echiniden-Gattungen. Verhandlungen der kaiserlich-königlichen geologischen Reichsanstalt, 6: 133–143.
|
| ○ |
Bodelle, J.; Campredon, R. & Lanteaume, M. (1968). Excursions dans les Alpes Maritimes et les Basses-Alpes. Colloque sur l’Éocène, 21–22 mai 1968. Bureau de Recherches géologiques et minières, Orléans, 73 pp.
|
| ○ |
Boschele, S.; Gatto, R.; Bernardi, M.; Bosellini, F.R. & Avanzini, M. (2016). Fossili cenozoici della Valsugana. Catalogo della “Collezione Boschele”, parte II. Studi Trentini di Scienze Naturali, 95: 53–102.
|
| ○ |
Bouillé, R. de (1873). Paléontologie de Biarritz et de quelques autres localités des Basses Pyrénées. Memoires de la Première Section, Congrès Scientifique de France, 39 Session 1, Pau, 427–468.
|
| ○ |
Bouillé, R. de (1876). Paléontologie de Biarritz et de quelques autres localités des Basses-Pyrenées. Imprimerie et Lithographie Veronese, Pau, 71 pp.
|
| ○ |
Boussac, J. (1908). Note sur la succession des faunes nummulitiques à Biarritz. Bulletin de la Société Géologique de France, 4ème série, 8(5): 237–255.
|
| ○ |
Boussac, J. (1911). Études stratigraphiques et paléontologiques sur le nummulitique de Biarritz. Annales Hébert. Annales de Stratigraphie et de Paléontologie du Laboratoire de Géologie de la Faculté des Sciences de l’Université de Paris, 5: 1–95.
https://doi.org/10.5962/bhl.title.139027 |
| ○ |
Brander, G. (1766). Fossilia Hantoniensia. Hampshire fossils. Fossilia Hantoniensia collecta, et in Musæo Britannico deposita, Londini, 43 pp. https://doi.org/10.5962/bhl.title.146466 |
| ○ |
Brongniart, A. (1823). Mémoire sur les terrains de sédiment supérieurs calcaréo-trappéens du Vicentin et sur quelques terrains d’Italie, de France, de l’Allemagne, etc. qui peuvent se rapporter à la même époque. Levrault, Paris, 84 pp. https://doi.org/10.5962/bhl.title.9097 |
| ○ |
Bronn, H.G. (1831). Uebersicht der fossilen Ueberreste in den tertiären subapenninischen Gebirgen. Ergebnisse meiner Naturhistorisch-ökonomischen Reisen, 2: 509–646.
|
| ○ |
Brzobohatý, R. & Nolf, D. (2011). Fish otoliths from the Middle Eocene (Bartonian) of Yebra de Basa, province of Huesca, Spain. Bulletin de l’Institut royal des Sciences naturelles de Belgique, Sciences de la Terre, 81: 279–295.
|
| ○ |
Buffrénil, V. de; Astibia, H.; Pereda Suberbiola, X.; Berreteaga, A. & Bardet, N. (2008). Variation in bone histology of middle Eocene sirenians from Western Europe. Geodiversitas, 30(2): 425–432.
|
| ○ |
Butts, S.H. (2014). Silicification. In: Reading and Writing of the Fossil Record: Preservational Pathways to Exceptional Fossilization (Laflamme, M.; Schiffbauer, J.D. & Darroch, S.A.F., Eds.). The Paleontological Society Papers, 20, The Paleontological Society Short Course, October 18, 2014, Cambridge, 15–33.
|
| ○ |
Calzada, S. & Astibia, H. (1996). Sobre Plicatula pamplonensis Carez: revisión. Bulletin de la Société d’Histoire naturelle de Toulouse, 132: 65–69.
|
| ○ |
Cappetta, H. (2012). Handbook of Paleoichthyology. Chondrichthyes, Mesozoic and Cenozoic Elasmobranchii: Teeth, Vol. 3E. Verlag Dr. Friedrich Pfeil, 512 pp.
|
| ○ |
Carez, L. (1881). Étude des Terrains crétacés et tertiaires du nord de l’Espagne. F. Savy, Paris, 327 pp. |
| ○ |
Carlsen, A.W. & Cuny, G. (2014). A study of the sharks and rays from the Lillebælt Clay (Early-Middle Eocene) of Denmark, and their palaeoecology. Bulletin of the Geological Society of Denmark, 62: 39–88.
|
| ○ |
Carpenter, P.H. (1884). Report upon the Crinoidea collected during the voyage of H.M.S. Challenger during the years 1873–1876. Part I. The Stalked Crinoids. Report on the Scientific Results of the Voyage of H.M.S. Challenger during the years 1873–76, Zoology, 11(32), i–xii, 1–442 pp. https://doi.org/10.5962/bhl.title.11805 |
| ○ |
Carrasco, J.F. (1994). El género Spondylus en el Eoceno del Noroeste de la Península Ibérica. Scripta Musei Geologici Seminarii Barcinonensis, 226: 3–24.
|
| ○ |
Carter, J.G.; Harries, P.J.; Malchus, N.; Sartori, A.F.; Anderson, L.C.; Bieler, R.; Bogan, A.E.; Coan, E.V.; Cope, J.C.W.; Cragg, S.M.; García-March, J.R.; Hylleberg, J.; Kelley, P.; Kleemann, K.; Kříž, J.; McRoberts, C.; Mikkelsen, P.M.; Pojeta, J. Jr.; Tëmkin, I.; Yancey, T. & Zieritz, A. (2012). Part N, Revised, Volume 1, Chapter 31: Illustrated Glossary of the Bivalvia. Treatise Online, 48: 1–209. https://doi.org/10.17161/to.v0i0.4322 |
| ○ |
Cascella, A. & Dinarès-Turell, J. (2009). Integrated calcareous nannofossil biostratigraphy and magnetostratigraphy from the uppermost marine Eocene deposits of the southeastern Pyrenean foreland basIn: evidences for marine Priabonian deposition. Geologica Acta, 7(1–2): 281–296.
|
| ○ |
Cherns, L.; Wheeley, J.R. & Wright, V.P. (2011). Taphonomic Bias in Shelly Faunas Through Time: Early Aragonitic Dissolution and Its Implications for the Fossil Record. In: Taphonomy, Second Edition - Process and Bias Through Time (Allison, P.A. & Bottjer, D.J., Eds.), Topics in Geobiology 32, Springer, Dordrecht. https://doi.org/10.1007/978-90-481-8643-3_3 |
| ○ |
Children, J.G. (1823). Lamarck’s Genera of Shells. The Quarterly Journal of Science, Literature and the Arts, 15: 23–52. |
| ○ |
Cossmann, M. (1898). Estudio de algunos moluscos eocenos del Pirineo catalán. Boletín de la Comisión del Mapa Geológico de España, 2(3): 167–198.
|
| ○ |
Cossmann, M. (1921). Synopsis illustré des mollusques de l’Éocène et de l’Oligocène de Aquitaine. Mémoires de la Société Géologique de France, Paléontologie, 55: 1–220.
|
| ○ |
Cossmann, M. & Pissarro, G. (1900). Faune éocénique du Cotentin (Mollusques). 1e article. Bulletin de la Société géologique de Normandie 19: 19–75. https://doi.org/10.5962/bhl.title.103164 |
| ○ |
Cotteau, G. (1863). Ëchinides fossiles des Pyrénées. Extrait du Congrès scientifique de France, 28e session, T. III, Chez J.-B. Baillière Libraire-Éditeur, Paris, Imp. et Lib. Maison Lafargue Coderc, Degréteau et Poujol, Succ., Bordeaux. https://doi.org/10.5962/bhl.title.11411 |
| ○ |
Cotteau, G. (1883). Échinides nouveaux ou peu connus 2. Bulletin de la Société zoologique de France, 8: 450–464. |
| ○ |
Courville, P.; Pacaud, J.-M.; Merle, D. & Lebrun, P. (2012). Le Lutétian de Damery (Marne, France): Géologie, environnements, associations de bivalves et gastéropodes. Fossiles, revue française de paléontologie, hors-série 3: 57–71.
|
| ○ |
D’Achiardi, A. (1867). Corallari fossili del terreno nummulitico dell’Alpi Venete, Catalogo delle specie e brevi note. Memorie della Società italiana di scienze naturali, 2: 1–18.
|
| ○ |
D’Achiardi, A. (1868). Corallari fossili del terreno nummulitico dell’Alpi Venete. Memorie della Società italiana di scienze naturali, parte 2, 4(1): 1–31.
|
| ○ |
D’Achiardi, A. (1875). Coralli Eocenici del Friuli. Atti della Società toscana di scienze naturali, residente in Pisa, 1: 1–102.
|
| ○ |
De Renzi, M. (1971). Las faunas de moluscos fósiles del Eoceno inferior del Prepirineo de Lérida. Tesis doctoral, Universitat de Barcelona, 502 pp.
|
| ○ |
De Renzi, M. (1996). La influencia de los factores tafonómicos y paleoecológicos en la distribución de los moluscos en el área tipo del Ilerdiense (Conca de Tremp, Cataluña, España). Revista Española de Paleontología, Nº extraordinario: 204–214.
|
| ○ |
Defrance, J.L.M. (1827). Serpule. In: Dictionnaire des sciences naturelles, dans lequel on traite me´thodiquement des diffe´rens êtres de la nature, conside´re´s soit en eux-mêmes, d’après l’e´tat actuel de nos connoissances, soit relativement à l’utilite´ qu’en peuvent retirer la me´decine, l’agriculture, le commerce et les artes. (Levrault, F.G., Ed.), Paris et Strasbourg, 48: 549–572.
|
| ○ |
Deshayes, G.-P. (1839-1853). Traité élémentaire de conchyliologie avec les applications de cette science à la géologie. Masson, Paris. https://doi.org/10.5962/bhl.title.125546 |
| ○ |
Deshayes, G.-P. (1860). Description des animaux sans vertèbres découverts dans le Bassin de Paris, 1, Atlas de 89 planches, J.-B. Baillère et Fills, Paris, 88 pp.
|
| ○ |
Desmarest, A.-G. (1817). Crustacés fossiles. In: Nouveau Dictionnaire d’Histoire naturelle, appliquée aux Arts, à l’Agriculture,
à l’Économie rurale et domestique, à la Médecine, etc., Société de Naturalistes et d’Agriculteurs, 7 (COR-CUN), Imprimerie d’Abel Lanoe, Deterville, Paris, 495–519.
|
| ○ |
Díaz-Berenguer, E.; Badiola, A.; Moreno-Azanza, M. & Canudo, J.I. (2018). First adequately-known quadrupedal sirenian from Eurasia (Eocene, Bay of Biscay, Huesca, northeastern Spain). Scientific Reports, 8: 5127. https://doi.org/10.1038/s41598-018-23355-w |
| ○ |
Dominici, S. & Kowalke, T. (2007). Depositional dynamics and the record of ecosystem stability: early Eocene coastal communities in the Pyrenean foreland, Spain. Palaios, 22: 268–284. https://doi.org/10.2110/palo.2005.p05-022r |
| ○ |
Dominici, S. & Kowalke, T. (2014). Early Eocene Cerithioidean gastropods of mangrove-fringed coasts (South-central Pyrenees, Spain). Bollettino della Società Paleontologica Italiana, 53(3): 137–162.
|
| ○ |
Doncieux, L. (1911). Catalogue descriptif des fossiles Nummulitiques de l’Aude et de Herault. Deuxième partie (fascicule II). Corbières septentrionals. Annales de l’Université de Lyon, série I Sciences, Médecine 30: 1–199.
|
| ○ |
Elorza, J. & Astibia, H. (2017). El anélido Rotularia spirulaea (Lamarck, 1818) (Polychaeta, Serpulidae) de las margas del Eoceno de la Cuenca de Pamplona (Navarra): microestructura, tafonomía y paleoecología. Spanish Journal of Palaeontology, 32(2): 343–366. https://doi.org/10.7203/sjp.32.2.17048 |
| ○ |
Elorza, J. & Astibia, H. (2018). Fosildiagénesis del anélido Rotularia spirulaea (Lamarck, 1818) (Polychaeta, Serpulidae) en el Eoceno del dominio pirenaico occidental. Spanish Journal of Palaeontology, 33(2): 299–320. https://doi.org/10.7203/sjp.33.2.13605 |
| ○ |
Esteban, F.J. (2006). Caracterización Microestructural y Cristalográfica de la Concha Prismatofoliada de Pectinoidea, Anomioidea y Ostreoidea (Pteriomorphia: Bivalvia). Implicaciones Evolutivas. Tesis doctoral, Universidad de Granada, 186 pp.
|
| ○ |
Fabiani, R. (1915). Monografia sui terreni terziari del Veneto. Parte 1: Il Paleogene del Veneto. Memorie dell’Istituto Geologico della R. Università di Padova, 3, XVI: 1–336.
|
| ○ |
Farrés, F. & Staid-Staadt, J.L. (1964). Las correlaciones faciales del Lediense y su fauna de moluscos en la comarca de Vich. Ausa, 5(48): 41–78.
|
| ○ |
Faura, M. (1926). El Tibidabo y Montserrat. Colección Barcelona y sus alrededores. Imprenta. V. Rico, Madrid, 42 pp. |
| ○ |
Felix, J. (1909). Über eine untertertiäre Korallenfauna aus der Gegend von Barcelona. Paleontographica, 56: 113–136. |
| ○ |
Fernández López, S. (1990). El significado de la autoctonía/aloctonía tafonómica. Comunicaciones de la Reunión Tafonomía y Fosilización, Madrid, 115–124.
|
| ○ |
Fernández López, S. (1991). Taphonomic concepts for a theoretical biochronology. Revista Española de Paleontología, 6(1): 37–49.
|
| ○ |
Fernández López, S. (2000). Temas de Tafonomía. Departamento de Paleontología, Universidad Complutense de Madrid, 167 pp. |
| ○ |
Fischer de Waldheim, G. (1835). Lettre à M. le Baron de Férussac sur quelques genres de coquilles du Muséum-Demidoff et en particulier sur quelques coquilles fossiles de la Crimée. Bulletin de la Société Impériale des Naturalistes de Moscou, 8: 99–123.
|
| ○ |
Fozy, I. & Szente, I. (2014). Fossils of the Carpathian Region. Indiana University Press, Bloomington and Indianapolis, 508 pp.
|
| ○ |
Fujiwara, S.-I.; Oji, T.; Tanaka, Y. & Kondo, Y. (2005). Relay Strategy and Adaptation to a Muddy Environment in Isselicrinus (Isselicrinidae: Crinoidea). Palaios, 20: 241–248.
|
| ○ |
Gemmellaro, G.G. (1868–1876). Studi paleontologici sulla fauna del calcare a Terebratula janitor del nord di Sicilia. Lao, Palermo, 56 pp.
|
| ○ |
Goldfuss, A. (1826–1844). Petrefacta Germaniae. Arnz & Co., Düsseldorf. |
| ○ |
Hernández Sampelayo, P. (1933). El Flysch en Yesa (Navarra). Notas y Comunicaciones del Instituto Geológico y Minero de España, 5: 74–76.
|
| ○ |
Herrmannsen A.N. (1846). Indicis generum malacozoorum primordia. Nomina subgenerum, generum, familiarum, tribum, ordinbum, classium; adjectis auctoribus, temporibus, locis systematicus atquae literariis, etymis, synonymis. Vol. 1. Fischer, Cassellis [Kassel], i-xxvii + 1–232. https://doi.org/10.5962/bhl.title.10147
|
| ○ |
Hess, H. & Messing, C.G. (2011). Revised, Crinoidea (3). In: Treatise on Invertebrate Paleontology, Part T, Echinodermata 2, (Selden, P.A., Ed.), The University of Kansas, Paleontological Institute, Lawrence, 1–261.
|
| ○ |
Hosgör, I. & Okan, Y. (2006). The annelid polychaete Rotularia spirulaea Lamarck, 1818 from the early Middle Eocene (middle-late Cuisian) of Çankırı Basin (Central Anatolia, Turkey). Journal of the Earth Sciences Application and Research Centre of Hacettepe University, 27: 173–179.
|
| ○ |
Jacquot, M.E. (1864). Description géologique des falaises de Biarritz, Bidart, Guétary et Saint-Jean-de-Luz. Actes de la Société Linnéenne de Bordeaux, 3ème série, 5: 1–58.
|
| ○ |
Jäger, M. (1983). Serpulidae (Polychaeta sedentaria) aus der norddeutschen höheren Oberkreide - Systematik, Stratigraphie,
Ökologie. Geologisches Jahrbuch, A68: 3–219.
|
| ○ |
Kirilova, L. & Georgiev, V. (2018). Fossil sea lilies of the genus Isselicrinus Rovereto, 1914 from the lower Oligocene, Eastern Rhodopes (Bulgaria). Geologica Balcanica, 47(1): 37–41.
|
| ○ |
Klikushin, V.G. (1982). Taxonomic survey of fossil isocrinids with a list of the species found in the USSR. Geobios, 15(3): 299–325. https://doi.org/10.1016/S0016-6995(82)80083-1 |
| ○ |
Kozlov, V.A. (2001). Sobre una nueva especie del tiburón Anomotodon de los sedimentos bartonienses de Mangyshlak. Materiales sobre estratigrafía y paleontología. Instituto de Geología y Geoquímica, Academia Rusa de Ciencias, Oficina de los Urales, Ekaterimburgo, 87–88 [en ruso].
|
| ○ |
Lamarck, J.-B. (1801). Systéme des animaux sans vertèbres. Deterville, Paris, 432 pp. |
| ○ |
Lamarck, J.-B. (1804). Suite des Mémoires. Sur les fossiles des environs de Paris (suite 3). Annales du Muséum d’Histoire naturelle, 4: 215–222.
|
| ○ |
Lamarck, J.-B. (1806). Suite des mémoires sur les fossiles des environs de Paris: Annales du Muséum National d’Histoire Naturelle, 7: 53–62.
|
| ○ |
Lamarck, J.-B. (1818). Histoire naturelle des animaux sans vertèbres, présentant les caractères généraux et particuliers de ces animaux, leur distribution, leurs classes, leurs familles, leurs genres et la citation des principales espèces qui s’y rapportent, précédée d’une introduction offrant la détermination des caractères essentiels de l’animal, sa distinction du végétal et des autres corps naturels, enfin, l’exposition des principes fondamentaux de la zoologie, 5. Déterville et Verdière, Paris, 612 pp.
|
| ○ |
Latasa, J.H. (1996). M. Ruiz de Gaona. Reseña humana de un escolapio. In: Homenaje a Máximo Ruiz de Gaona, naturalista y paleontólogo (1917–1971) (Astibia, H.; del Valle de Lersundi, J.; Murelaga, X. & Serra-Kiel, J., Coords.), Príncipe de Viana (Suplemento de Ciencias), 14, Gobierno de Navarra, Pamplona, 65–76.
|
| ○ |
Leymerie, A. (1881). Description géologique et paléontologique des Pyrénnées de l’Haute Garonne. Edición privada, Toulouse, 1010 pp.
|
| ○ |
Leymerie, A. & Cotteau, G.(1856). Catalogue des Échinides fossiles des Pyrénnées. Bulletin de la Société géologique de France, Série 2, 13: 319–355.
|
| ○ |
Leymerie, M.A. (1846). Mémoire sur le terrain à Nummulites (Epicrétacé) des Corbières et de la Montagne Noire. Mémoires de la Société Géologique de France, 2ème série, 8: 337–373.
|
| ○ |
Linnaeus, C. (1758). Systema Naturae per Regna Tria Naturae, Secundum Classes, Ordines, Genera, Species, cum Characteribus, Differentiis, Synonymis, Locis. Tenth edition. Laurentii Salvii, Holmiae, Stockholm, 824 pp. https://doi.org/10.5962/bhl.title.542 |
| ○ |
López-Horgue, M.A. & Bodego, A. (2017). Mesozoic and Cenozoic decapod crustaceans from the Basque-Cantabrian basin (Western Pyrenees): new occurrences and faunal turnovers in the context of basin evolution. Bulletin de la Société géologique de France, 188, 14. https://doi.org/10.1051/bsgf/2017180 |
| ○ |
Loriol, P. de (1880). Monographie des échinides contenus dans les couches nummulitiques de l’Egypte. Mémoire de la Société de Physique et d’Histoire naturelle de Genève, 27(1): 57–148.
|
| ○ |
Malubián, R. & Olivero, E.B. (2005). Shallow-water lat middle Eocene crinoids from Tierra del Fuego: a new southern record of a retrograde community structure. Scientia Marina, 69 (Suppl. 2): 349–353. https://doi.org/10.3989/scimar.2005.69s2349 |
| ○ |
Mannering, A.A. & Hiller, N. (2008). An early Cenozoic neoselachian shark fauna from the Southwest Pacific. Palaeontology, 51(6): 1341–1365. https://doi.org/10.1111/j.1475-4983.2008.00812.x |
| ○ |
Marquina, F. (1908). Descripción geológica de Navarra. In: Geografía general del País Vasco-Navarro. Altadill J., Provincia de Navarra (Carreras y Candi F., Ed.), Editorial Alberto Martín, Barcelona, 215–287.
|
| ○ |
Martinius, A.W. (1991). Growth rates and population dynamics in Crassostrea cf. rarilamella from the Lower Eocene Roda Formation (southern Pyrenees, Spain). Geologie en Mijnbouw, 70: 59–73.
|
| ○ |
Mathelin, J.C. & Sztràkos, K. (1993). L’Éocène de Biarritz (Pyrénées-Atlantiques, SW France). Stratigraphie et paléoenvironnement. Monographie des foraminifères. Cahiers de Micropaléontologie, nouvelle série, 8(1–2): 5–85.
|
| ○ |
Matteucci, R. & Russo, A. (2005). The Middle Eocene siliceous sponges from Val di Chiampo (Lessini Mountains, northern Italy). Annali dell’Università di Ferrara, Sezione di Museologia Scientifica e Naturalistica special: 51–62.
|
| ○ |
Melleville, M. (1843). Mémoire sur les sables tertiaires inférieurs du Bassin de Paris, avec la description de 78 espèces de coquilles fossiles inédites de ce terrain. Annales des Sciences Géologiques, 2: 1–75.
|
| ○ |
Mendizábal, J. & Ruiz de Gaona, M. (1949). Hoja geológica num. 141 (Pamplona). Mapa Geológico de España E. 1:50.000. Instituto Geológico y Minero de España. Madrid.
|
| ○ |
Merle, D. (2008). Le Lutétien: un exemple de point-chaud de la paléobiodiversité. In: Stratotype Lutétien. Muséum national d’Histoire naturelle, Paris, Biotope, Mèze (Merle, D., Coord.), Collection Patrimoine géologique 1, BRGM, Orléans, 174–181.
|
| ○ |
Michelin, H. (1841). Iconographie zoophytologique. Description par localités et terrains des Polypiers fossiles de France et pays environnants. P. Bertrand, 1840–1847, Paris, 40 pp. https://doi.org/10.5962/bhl.title.11504 |
| ○ |
Mikuž, V. (2008). The serpulid worm Rotularia spirulaea from Eocene beds near Gracisce in Istria, Croatia. Geologija, 51: 161–168. https://doi.org/10.5474/geologija.2008.017 |
| ○ |
Miller, J.S. (1821). A Natural History of the Crinoidea of Lily-shaped Animals, with Observations on the Genera Asterias, Euryale, Comatula, and Marsupites. Bryan & Co, Bristol, 150 pp. https://doi.org/10.5962/bhl.title.32130 https://doi.org/10.5962/bhl.title.11565 |
| ○ |
Montfort, D. de (1810). Conchyliologie systématique et classification méthodique des coquilles. Coquilles univalves, non cloisonnées, tome 2, Schoell, Paris, 676 pp.
|
| ○ |
O’Gorman, G. & Cossmann, M. (1923). Le gisement cuisien de Gan (Basses-Pyrénées). Chez M. Cossmann, Pau, 188 pp. |
| ○ |
Oppenheim, P. (1901). Über einige alttertiäre Faunen der Österreichisch-Ungarischen Monarchie. Beiträge zur Paläontologie und Geologie Österreich-Ungarns und des Orients, 13: 145–277.
|
| ○ |
Oppenheim, P. (1903). Zur Kenntnis alttertiärer Faunen in Ägypten. Bivalven. Palaeontographica, Teil 1, 30: 1–164. |
| ○ |
Oppenheim, P. (1910). Bemerkungen zu Prof. Johannes Felix: “Über eine untertertiäre Korallenfauna aus der Gegend von Barcelona”. Zeitschrift der Deutschen Geologischen Gesellschaft, 62 (2): 129–141.
|
| ○ |
Oppenheim, P. (1911). Weitere Notizen zur alttertiären Korallenfauna von Barcelona. Briefliche Mitteilungen, 23: 329–356. |
| ○ |
Orbigny, A. d’ (1847). Considérations zoologiques et géologiques sur les brachiopodes ou palliobranches, parts 1–2. Comptes Rendus Hebdomadaires des Séances de l’Académie des Sciences, Paris, 2 ème série, 25(5): 193–195; 25(7): 266–269.
|
| ○ |
Orbigny, A. d’ (1849). Description de quelques genres nouveaux de Mollusques bryozoaires. Revue et Magasin de zoologie pure et appliquée 2e série, 1: 499–504.
|
| ○ |
Orbigny, A. d’(1850). Prodrome de Paléontologie stratigraphique universelle des animaux mollusques et rayonnés 2. Masson, Paris, 427 pp. https://doi.org/10.5962/bhl.title.62810 |
| ○ |
Payros, A. (1997). El Eoceno de la cuenca de Pamplona: estratigrafía, facies y evolución paleogeográfica. Tesis doctoral, Universidad del País Vasco, 403 pp.
|
| ○ |
Payros, A., Dinarès-Turell, J., Bernaola, G., Orue-Etxebarria, X., Apellaniz, E. & Tosquella, J. (2011). On the age of the Early/Middle Eocene boundary and other related events: cyclostratigraphic refinements from the Pyrenean Otsakar section and the Lutetian GSSP. Geological Magazine, 148: 442–460. https://doi.org/10.1017/S0016756810000890 |
| ○ |
Payros, A.; Astibia, H.; Cearreta, A.; Pereda-Suberbiola, X.; Murelaga, X. & Badiola, A. (2000). The Upper Eocene South Pyrenean Coastal Deposits (Liédena Sandstone, Navarre): Sedimentary Facies, Benthic Foraminifera and Avian Ichnology. Facies, 42: 107–132.
https://doi.org/10.1007/BF02562569 |
| ○ |
Payros, A.; Orue-Etxebarria, X. & Pujalte, V. (2006). Covarying sedimentary and biotic fluctuations in Lower-Middle Eocene Pyrenean deep-sea deposits: palaeoenvironmental implications. Palaeogeography, Palaeoclimatology, Palaeoecology, 234: 258–276.
https://doi.org/10.1016/j.palaeo.2005.10.013 |
| ○ |
Payros, A.; Pujalte, V. & Orue-Etxebarria, X. (1999). The South Pyrenean Eocene carbonate megabreccias revisited: new interpretation based on evidence from the Pamplona Basin. Sedimentary Geology, 125: 165–194. https://doi.org/10.1016/S0037-0738(99)00004-4 |
| ○ |
Payros, A.; Pujalte, V. & Orue-Etxebarria, X. (2007). A point-sourced calciclastic submarine fan complex (Eocene Anotz Formation, western Pyrenees): facies architecture, evolution and controlling factors. Sedimentology, 54: 137–168. https://doi.org/10.1111/j.1365-3091.2006.00823.x |
| ○ |
Payros, A.; Pujalte, V.; Tosquella, J. & Orue-Etxebarria, X. (2010). The Eocene storm-dominated foralgal ramp of the western Pyrenees (Urbasa-Andia Formation): An analogue of future shallow-marine carbonate systems? Sedimentary Geology, 228: 184–204.
https://doi.org/10.1016/j.sedgeo.2010.04.010 |
| ○ |
Pelc, A.; Hałas, S. & Niedźwiedzki, R. (2011). Oxygen isotope analysis of shark teeth phosphates from Bartonian (Eocene) deposits in Mangyshlak peninsula, Kazakhstan. Mineralogia, 42(1): 53–61. https://doi.org/10.2478/v10002-011-0006-7 |
| ○ |
Philippi, R.A. (1846). Verzeichniss der in der Gegend von Magdeburg aufgefundenen Tertiärversteinerungen. Palaeontographica, 1(1): 42–44.
|
| ○ |
Piccoli, G. & Mocellin, L.G. (1962). Studi sulla macrofauna Priaboniana di Priabona (Prealpi venete). Memorie degli Istituti di Geololia e Mineralogia dell’Università di Padova, 23: 1–120.
|
| ○ |
Pisera, A. & Busquets, P. (2002). Eocene siliceous sponges from the Ebro Basin (Catalonia, Spain). Les éponges siliceuses de l’Éocène du Basin de l’Èbre (Catalogne, Espagne). Geobios, 35: 321–346. https://doi.org/10.1016/S0016-6995(02)00030-X |
| ○ |
Plaziat, J.C. (1981). Late Cretaceous to Late Eocene paleogeographic evolution of southwest Europe. Palaeogeography, Palaeoclimatolology, Palaeoecology, 36: 263–320. https://doi.org/10.1016/0031-0182(81)90110-3 |
| ○ |
Puigdefábregas, C. (1975). La sedimentación molásica en la cuenca de Jaca. Pirineos, 104: 1–188. |
| ○ |
Pujalte, V.; Baceta, J.I. & Payros, A. (2002). Tertiary: Western Pyrenees and Basque-Cantabrian region. In: The Geology of Spain (Gibbons, W. & Moreno, T., Eds.), Geological Society of London, London, 293–301.
|
| ○ |
Rasmussen, H.W. (1954). Cretaceous Crinoidea. Second preliminary report. Meddelelser fra Dansk Geologisk Forening. København, 12: 553–555,
|
| ○ |
Rasmussen, H.W. (1972). Lower Tertiary Crinoidea, Asteroidea and Ophiuroidea from Northern Europe and Greenland. Biologiske Skrifter udgivet af det Kgl. Danske Videnskabernes Selskab, 19: 1–83.
|
| ○ |
Reguant, S. (1993). Semi-multilamellar growth in Reptomulticava alhamensis, a new cyclostome (Bryozoa) species from the Tortonian if alhama de Granada (S. Spain). Revista Española de Paleontología, 8(2): 147–152.
|
| ○ |
Reinecke, T.; Stapf, H. & Raisch, M. (2001). Die Selachier und Chimären des Unteren Meeressandes und Schleichsandes im Mainzer Becken (Alzey- und Stadecken-Formation, Rupelium, Unteres Oligozän). Palaeontos, 1: 1–73.
|
| ○ |
Reish, D.J. (1968). The Polychaetous Annelids of the Marshall Islands. Pacific Science, 22(2): 208–231. |
| ○ |
Reuss, A. E. (1874). Paläontologische Studien über die älteren Tertiärschichten der Alpen. III Abtheilung. Die Fossilen Anthozoen der Schichtengruppe von S. Giovanni Ilarione und von Roncà. Nachträge zu den ersten zwei abtheilungen. Schlussbemerkungen. Allgemeines Namensregister. Denkschriften der kaiserlichen Akademie der Wissenschaften, Mathematisch-Naturwissenschaftliche Classe, 33: 1–60.
|
| ○ |
Röding, P.F. (1798). Museum Boltenianum sive Catalogus cimeliorum e tribus regnis naturae quae olim collegerat Joa. Fried Bolte, Pars Secunda. Typis Johan. Christi. Trappii, Hamburgi, 199 pp.
|
| ○ |
Rouault, A. (1850). Description des fossiles du terrain éocène des environs de Pau. Mémoires de la Société géologique de France, 2(3): 457–502.
|
| ○ |
Roux, M.; Cahuzac, B. & Sztràkos, K. (2006). Les paléoenvironnements éocènes à crinoïdes pédonculés des marnes de Miretrain (Angoumé, SW France): interprétations paléobathymétriques. Comptes Rendus Géoscience, 338(4): 262–271. https://doi.org/10.1016/j.crte.2006.01.001 |
| ○ |
Rovereto, G. (1914). Nuovi studi sulla stratigrafia e sulla fauna dell’Oligocene ligure. Olivieri E. & C., Società Tipo-Lito-grafica Ligure, Genova, 179 pp |
| ○ |
Ruiz de Gaona, M. (1947a). El Bartoniense de la Cuenca de Pamplona. Notas y Comunicaciones del Instituto Geológico y Minero de España, 17: 155–165.
|
| ○ |
Ruiz de Gaona, M. (1947b). Sobre el Eoceno de Urbasa a lo largo de la carretera provincial de Olazagutía a Estella. Estudios Geológicos, 5: 179–208.
|
| ○ |
Ruiz de Gaona, M. & Colom, G. (1950). Estudios sobre las sinecias de los foraminíferos eocénicos de la vertiente meridional del Pirineo (Cataluña-Vizcaya). Estudios Geológicos, 12: 293–434.
|
| ○ |
Russo, A. (1979). Studio monografico sui coralli dell’Eocene di Possagno (Treviso, Italia). Atti e memorie della Accademia nazionale di scienze, lettere e arti, Modena, 6, 21:1–87.
|
| ○ |
Sacco, F. (1895). I Molluschi dei Terreni Terziarii del Piemonte e della Liguria. Parte XVII. (Cerithiidae, Triforidae, Cerithiopsidae e Diastomidae). Memorie della Reale Accademia delle Scienze di Torino, Serie 2, 83 pp.
|
| ○ |
Salazar, Á.; Carcavilla, L.; Díaz-Martínez, E.; García-Cortés, Á. & Vegas, J. (2017). Yacimientos paleontológicos desaparecidos ¿Por qué inventariarlos? Paleontological sites disappeared. Why inventory them? In: Patrimonio geológico, gestionando la parte abiótica del patrimonio natural (Carcavilla, L.; Duque-Macías, J.; Giménez, J.; Hilario, A.; Monge-Ganuzas, M.; Vegas, J. & Rodríguez, A., Eds.), Cuadernos del Museo Geominero, 21, Instituto Geológico y Minero de España, Madrid, 33–38. |
| ○ |
Sanders, M.T.; Merle, D. & Villier, L. (2015). The molluscs of the “Falunière” of Grignon (Middle Lutetian, Yvelines, France): quantification of lithification bias and its impact on the biodiversity assessment of the Middle Eocene of Western Europe. Geodiversitas, 37(3): 345–365. https://doi.org/10.5252/g2015n3a4 |
| ○ |
Savazzi, E. (1995). Morphology and mode of life of the polychaete Rotularia. Paläontologische Zeitschrift, 69: 73–85. https://doi.org/10.1007/BF02985975 |
| ○ |
Schindler, T.; Nungesser, K.; Müller, A. & Grimm, K.I. (2009). Die Alzey-Formation der klassischen Lokalität Welschberg bei Waldböckelheim (Rupelium, Oligozän, Mainzer Becken) - Ergebnisse neuer Grabungen. Jahresberichte und Mitteilungen des Oberrheinischen Geologischen Vereins, Band 91: 37–87. https://doi.org/10.1127/jmogv/91/2009/37 |
| ○ |
Schmidt, F.C. (1818). Versuch über die beste Einrichtung zur Aufstellung: Behandlung und Aufbewahrung der verschiedenen Naturkörper und gegenṣtände der Kunst, vorzüglich der Conchylien-Sammlungen, nebst kurzer Beurtheilung der conchyliologischen Systeme und Schriften und einer tabellarischen Zusammenstellung und Vergleichung der sechs besten und neuesten conchyliologischen Systeme. Gotha, J. Perth, 252 pp.
|
| ○ |
Schnetler, K.I. & Heilmann-Clausen, C. (2011). The molluscan fauna of the Eocene Lillebælt Clay, Denmark. Cainozoic Research, 8(1–2): 41–99.
|
| ○ |
Seilacher, A.; Olivero, E.B.; Butts, S.H. & Jäger, M. (2008). Soft-bottom tube worms: from irregular to programmed shell growth. Lethaia, 41: 349–365. https://doi.org/10.1111/j.1502-3931.2008.00092.x |
| ○ |
Serra-Kiel, J.; Hottinger, L.; Caus, E.; Drobne, K.; Ferrandez, C.; Jauhri, A.K.; Less, G.; Pavlovec, R.; Pignatti, J.; Samso, J.M.; Schaub, H.; Sirel, E.; Strougo, A.; Tambareau, Y.; Tosquella, J. & Zakrevskaya, E. (1998). Larger foraminiferal biostratigraphy of the Tethyan Paleocene and Eocene. Bulletin de la Société géologique de France, 169: 281–299.
|
| ○ |
Serra-Kiel, J.; Travé, A.; Mató, E.; Saula, E; Ferràndez-Cañadell, C.; Busquets, P.; Tosquella, J. & Vergés, J. (2003). Marine and Transitional Middle/Upper Eocene Units of the Southeastern Pyrenean Foreland Basin (NE Spain). Geologica Acta, 1(2): 177–200.
|
| ○ |
Sowerby, J. (1812–1815). The mineral conchology of Great Britain; or coloured figures and descriptions of those remains of testaceous animals or shells, which have been preserved at various times and depths in the earth. Edición privada, Londres.
https://doi.org/10.5962/bhl.title.14408 |
| ○ |
Taylor, P.D. & Wilson, M.A. (2003). Palaeoecology and evolution of marine hard substrate communities. Earth-Science Reviews, 62: 1–103. https://doi.org/10.1016/S0012-8252(02)00131-9 |
| ○ |
Taylor, P.D.; Lombardi, Ch. & Cocito, S. (2015). Biomineralization in bryozoans: present, past and future. Biological Reviews, 90: 1118–1150. https://doi.org/10.1111/brv.12148 |
| ○ |
Tournouër, R. (1872). Note sur les fossiles tertiaires des Basses-Alpes, recueillis par M. Garnier. Bulletin de la Société Géologique de France, 2 ème série, 29: 492–527.
|
| ○ |
Tournouër, R. (1874). Sur les Échinides nummulitiques de Biarritz et rectifications de noms spécifiques. Bulletin de la Société Géologique de France, 3 ème série, 2: 527–528.
|
| ○ |
Vía, L. (1959). Decápodos fósiles del Eoceno español. Boletín del Instituto Geológico y Mineros de España, 70: 331–402. |
| ○ |
Villalta Comella, F. de (1956). Los moluscos fósiles del Eoceno pirenaico. Boletín del Instituto Geológico y Minero de España, 67: 119–235.
|
| ○ |
Vinassa de Regny, P. (1897). Synopsis dei molluschi terziari delle Alpi venete. (Continuazione e fine della parte prima): IV. Zovencedo (Tufo glauconitico). V. Monte Pulli, Caldiero, Bolca ecc. Parte seconda: Strati oligocenici. VI. Via degli Orti, Valle Orgagna, Priabona ecc. Palaeontographia Italica, 3: 145–200.
|
| ○ |
Wade, B.S.; Pearson, P.N.; Berggren, W.A. & Pälike, H. (2011). Review and revision of Cenozoic tropical planktonic foraminiferal biostratigraphy and calibration to the geomagnetic polarity and astronomical time scale. Earth-Science Reviews, 104: 111–142.
https://doi.org/10.1016/j.earscirev.2010.09.003 |
| ○ |
Waller, T.R. (2012). Morphology, phylogeny, and sistematic revision of genera in the Dimyidae (Mollusca, Bivalvia, Pteriomorphia). Journal of Paleontology, 86(5): 829–851. https://doi.org/10.1666/12-004.1 |
| ○ |
Whittle, R.J.; Hunter, A.W.; Cantrill, D.J. & McNamara, K.J. (2018). Globally discordant Isocrinida (Crinoidea) migration confirms asynchronous Marine Mesozoic Revolution. Communications Biology, 1: 46. https://doi.org/10.1038/s42003-018-0048-0 |
| ○ |
Wright, P.; Cherns, L. & Hodges, P. (2003). Missing molluscs: Field testing taphonomic loss in the Mesozoic through early large-scale aragonite dissolution. Geology, 31(3): 211–214. https://doi.org/10.1130/0091-7613(2003)031<0211:MMFTTL>2.0.CO;2 |
| ○ |
Wrigley, A. (1951). Some Eocene serpulids. Proceedings of the Geologists Association of the United Kingdom, 62: 177–197. |
| ○ |
Zamora, S. (2018). Crinoideos fósiles de Aragón. In: Fósiles. Nuevos hallazgos paleontológicos en Aragón (Zamora, S., Ed.), Institución Fernando el Católico, Zaragoza, 22–47.
|
| ○ |
Zamora, S.; Aurell, M.; Veitch, M.; Saulsbury, J.; López-Horgue, M.A.; Ferratges F.A.; Arz, J.A., & Baumiller, T.K. (2018). Environmental distribution of post-Palaeozoic crinoids from the Iberian and south-Pyrenean basins, NE Spain. Acta Palaeontologica Polonica, 63(4): 779–794. https://doi.org/10.4202/app.00520.2018 |
| ○ |
Zavarei, A. (1973). Monographie des Spondylidae (Lamellibranches) actuels et fossils. Tesis doctoral, Université de Paris-Sud, Centre d’Orsay, Série A (1147), 233 pp.
|
| ○ |
Zhelezko, V.I. & Kozlov, V. (1999). Elasmobranchii y bioestratigrafía del Paleógeno de los Trans-Urales y Asia Central. Materiales sobre estratigrafía y paleontología de los Urales, vol. 3. Instituto de Geología y Geoquímica, Academia Rusa de Ciencias, Oficina de los Urales, Ekaterimburgo, 324 pp. [en ruso].
|