Valorisation of palaeontological collections: nomination of a lectotype for Conohyus simorrensis (Lartet, 1851), Villefranche d’Astarac, France, and description of a new genus of tetraconodont
Puesta en valor de colecciones paleontológicas: designación de
un lectotipo para Conohyus simorrensis (Lartet, 1851), Villefranche
d’Astarac (Francia) y descripción de un nuevo género de tetraconodonto
M. Pickford1, Y. Laurent2
1Department Histoire de la Terre, UMR 7207 du CNRS, 8 rue Buffon, 75005, Paris, France. Email: pickford@mnhn.fr
2Muséum d’Histoire Naturelle, 35 allées Jules Guesdes, 31000 Toulouse, France
| |
ABSTRACT
The type species of the suid genus Conohyus Pilgrim (1925) is Sus simorrensis Lartet (1851) by original designation, but, as was usual at the time of the creation of the species in 1851, no holotype
was nominated. Allusion was made by Lartet (1851) to upper and lower canines and some molars from two localities, Simorre
and Villefranche d’Astarac, France. The canines belong to Listriodon splendens, a highly lophodont suid, but the molars were described as being bunodont. Until now a lectotype has not been nominated,
partly because the original collections of Lartet (1851) were thought to be lost, but also because the diagnosis of the molars
as bunodont was insufficient for distinguishing the species from a host of other suids. A suggestion was made to nominate
a mandible from Le Fousseret as lectotype, but it was never formalised, and later it was proposed that a bunodont tooth from
Simorre collected during the 1960s should be selected, but none was nominated.
We here report the re-discovery of the collection of suids examined by Lartet (1851) and select a lectotype for the species
Sus simorrensis. We provide detailed descriptions and illustrations of all the tetraconodont material from Villefranche d’Astarac, Simorre
(Rajegats) and Le Fousseret, and discuss the relationships of the fossils to other tetraconodont samples from Europe.
There are two taxa of bunodont tetraconodonts at Villefranche d’Astarac and Simorre, one of which represents a new genus,
which is defined and described.
Keywords: Tetraconodontinae;
Miocene;
France;
Villefranche d’Astarac;
Conohyus simorrensis;
lectotype;
new genus
|
| |
RESUMEN
Por designación original, la especie tipo del género de suidos Conohyus Pilgrim (1925) es Sus simorrensis Lartet (1851), pero como era habitual en el momento de la creación de la especie en 1851, no se designó el holotipo. Lartet
(1851) aludió a los caninos superiores e inferiores y algunos molares de dos localidades: Simorre y Villefranche d’Astarac,
Francia. Los caninos pertenecen a Listriodon splendens un suido extremadamente lofodonto, pero los molares se describieron como bunodontos. Hasta ahora no se ha designado el lectotipo,
en parte porque se pensaba que las colecciones originales de Lartet (1851) se habían perdido, pero también porque el diagnóstico
de los molares como bunodontos era insuficiente para distinguir la especie a partir de una serie de otros suidos. Aunque se
propuso nominar una mandíbula de Le Fousseret como lectotipo, nunca se formalizó, y más tarde se propuso que debía seleccionarse
un diente bunodonto del Simorre obtenido durante la década de 1960, pero tampoco fue designado.
En este trabajo se reporta el re-descubrimiento de la colección de suidos examinados por Lartet (1851) y se selecciona un
lectotipo para la especie Sus simorrensis. Ofrecemos descripciones detalladas e ilustraciones de todo el material tetraconodonto de Villefranche d’Astarac, Simorre
(Rajegats) y Le Fousseret, y se discuten las relaciones de los fósiles con otras muestras de tetraconodontos de Europa.
Hay dos taxones de tetraconodontos bunodontos en Villefranche d’ Astarac y Simorre, uno de los cuales representa un nuevo
género, que se define y describe.
Palabras clave: Tetraconodontinae;
Mioceno;
Francia;
Villefranche d’Astarac;
Conohyus simorrensis;
lectotipo;
nuevo género
|
IntroductionTOP
As was the custom during the mid-19th century, authors generally did not nominate holotypes for the species that they named, and Lartet (1851) was no exception.
In cases where only a single specimen is listed it is automatically the holotype, but in cases where several specimens are
described, a lectotype must be nominated, preferably from the original hypodigm of the creating author. In the case of Sus simorrensis, Lartet (1851) mentioned upper and lower canines from Simorre which were figured by Blainville (1847) and « molaires fort semblables à celles du Cochon, si ce n’est que leur couronne est moins parsemée de tubercules intermédiaires
aux mamelons principaux ». Lartet (1851) wrote that Sus simorrensis was collected from Simorre and Villefranche d’Astarac, but he reported that it was absent at Sansan.
The absence of a holotype for Sus simorrensis has been a stumbling block in tetraconodont taxonomy for well over a century and a half (Stehlin, 1899–1900; Hünermann, 1968; Chen, 1984; Van der Made, 1998). The usual comparative basis for Conohyus has comprised a mixture of at least three suid species (Sus simorrensis, Sus doati and Chaeropotamus steinheimensis).
Lartet’s (1851) hypodigm of Conohyus simorrensis includes remains of three taxa – Listriodon splendens, which is a suid with lophodont molars, and two taxa with bunodont Sus-like molars but divergent premolar morphology. The upper and lower canines from Simorre mentioned by Lartet (1851) were illustrated
by de Blainville (1847) and have been positively identified in the collections; they belong to Listriodon splendens. The bunodont suid specimens from Villefranche d’Astarac were thought to be lost (Van der Made, 1989) and as a consequence a proposal was made to nominate a mandible (MNHN HGP 16) from Le Fousseret, France, as a neotype.
Indeed, during the 1960’s, casts of this mandible which contains p/4-m/3 were distributed to various museums in Europe mis-labelled
“holotype of Conohyus simorrensis, Simorre”. The cast of a left mandible from Le Fousseret containing p/3-m/2 kept in Bologna (un-catalogued display cast of
MHNT PAL.2012.0.206) on which Stehlin (1899–1900) based his comparisons, is part of the same individual. Thus casts of two
fossils have acted as reference specimens for Conohyus simorrensis, and neither of them came from the type locality. The potential for confusion is obvious. Given the complexity of the local
stratigraphy in Haute Garonne, France (Richard, 1946) nominating a lectotype from among fossils from a site such as Le Fousseret which is 32 km from Simorre seems a hazardous
proposal.
In 1925, Pilgrim erected the genus Conohyus for medium sized tetracondont suids from the Miocene of Eurasia and nominated Sus simorrensis as the type species. However, because no type specimen had been proposed for Sus simorrensis Lartet, 1851, the genus was poorly defined, even though most authors have accepted Pilgrim’s (1925, 1926) opinion that Conohyus simorrensis was a well-defined taxon. Pilgrim’s (1925, 1926) concept of Conohyus simorrensis was based on Stehlin’s (1899–1900) treatment of Hyotherium simorrense, which was in its turn based on Fraas’ (1870) description of Chaeropotamus steinheimensis from Steinheim, Germany, Hofmann’s (1893) descriptions of the fossils from Göriach, Austria, and his own description of lower premolars from Tutzing, Germany. However,
restudy of the fossils indicates that at least three taxa are represented in Stehlin’s (1899–1900) concept of the species.
This makes Stehlin’s concept of the species a chimera, and means that hasty nomination of a lectotype could have deleterious
effects on the nomenclature of these suids.
Vitry (1857) in the Journal de Toulouse, dated 16th May, 1857, records that M. Leymerie mentioned the discovery of fossils by M. Abadie,
Pharmacist at Le Fousseret, and proposed his name for a medal of the Académie Impériale des Sciences, Inscriptions et Belles-Lettres
de Toulouse. Among the fossils found by M. Abadie, are two mandibles that were intially part of the collections of M. Leymerie,
but which were eventually donated to the MNHN Paris (MNHN HGP 16), and the MHN Toulouse (MHNT PAL 2012.0.206). This declaration
indicates that the fossil mandibles from Le Fousseret were likely collected after Lartet’s (1851) description of Sus simorrensis, and could not have influenced his decision to erect the species Sus simorrensis.
A cast of the teeth of the suid mandible from Le Fousseret (MHNT PAL 2012.0.206) housed in Toulouse which was donated to the
University of Bologna, Italy, was mentioned by Stehlin (1899–1900), Van der Made (1998) and Van der Made & Morales (2003). There is a problem with the description of the Fousseret cast in Bologna, in that it was initially reported to contain
the p/2-m/2 (Stehlin, 1899–1900, p. 45), whereas the original fossil from Le Fousseret kept in Toulouse (MHNT PAL 2012.0.206)
contains only the left p/3-m/2, and the opposite jaw, kept in Paris (MNHN HGP 16) contains the right p/4-m/3. The contradiction
arises from the fact that Stehlin (1899–1900) counted the premolars from the rear towards the front, thus the p/4 was called
by him the p/1, and the p/3 was called p/2. Stehlin (1899–1900, p. 140 footnote) observed the original fossil from Le Fousseret
at Toulouse but did not find the series of specimens of Conohyus simorrensis from Simorre and Villafranche d’Astarac in the collections.
Van der Made (1989) wrote that a mandible from Le Fousseret, France, housed in the Muséum National d’Histoire Naturelle, Paris, will be nominated
as a neotype by Ginsburg (paper never published). However, Le Fousseret is 32 km from Villefranche d’Astarac and Simorre,
and the premolars in the mandible are larger than those in the material from Villafranche d’Astarac (Stehlin, 1899–1900; Hünermann, 1968). Casts of the MNHN HGP 16 mandible were distributed to other museums during the 1960’s, with the erroneous information that
it was from Simorre, and furthermore that it was the holotype (see for example, the cast housed in the Hessisches Landesmuseum,
Darmstadt (HLMD Fr 1b)).
Van der Made (1998) discussed the issue of a neotype for Conohyus simorrensis, but suggested that for the sake of stability, instead of the mandible from Le Fousseret previously proposed by him (Van der Made, 1989) a bunodont specimen from excavations made by Ginsburg at Simorre should be selected. The disadvantage of this suggestion
is fourfold: 1) the Ginsburg material consists of isolated teeth collected more than a Century after the creation of the species,
2) the type locality turns out to be Villefranche d’Astarac, and not Simorre, 3) whilst most of the suid teeth from Simorre
and Villefranche d’Astarac have comparable dimensions, the upper third molars from Simorre are significantly smaller than
the one from Villefranche d’Astarac, and 4) there are two bunodont suids at Villefranche d’Astarac, one with sectorial p/2
at the same level as the p/3, the other with bunodont p/2 vertically offset beneath the level of the p/3.
Recent valorisation of the collections at the Muséum d’Histoire Naturelle, Toulouse, has led to the rediscovery of several
specimens labelled on the fossils themselves with the words “Sus simorrensis Villefranche d’Astarac” in the handwriting of E. Lartet. In addition they have paper labels stuck to them with the words
“COLLECTIONS Edouard et Louis LARTET” and “N° …. du Cat. Spécial” printed in red ink, typical of the collections of E. Lartet.
There can be little doubt that this material is what Lartet (1851) had in mind when he wrote the paper in which he erected the species Sus simorrensis. The left mandible with p/4-m/3 has the letter “i” written on it in red paint beneath the m/3 on the lingual side, but the
significance of this is not known (it could mean “illustré”).
The aim of this paper is to describe and interpret the re-discovered suid fossils from Villefranche d’Astarac and to select
a lectotype for the species Sus simorrensis. A second aim is to discuss the consequences of the discovery of these fossils on the taxonomy of tetraconodonts, one of
which is the recognition of a new genus of tetraconodont.
Materials and MethodsTOP
The fossils described herein are curated at the Muséum d’Histoire Naturelle, Toulouse, and the Muséum National d’Histoire
Naturelle, Paris. An un-catalogued cast of a left mandible, from Le Fousseret, the original of which is housed at the Natural
History Museum Toulouse (MHNT PAL.2012.0.206) is in the university collection in Bologna, Italy. Although the latter cast
is quite poor, it is historically an important specimen because it contributed to the formation of Stehlin’s (1899–1900) concept
of the species. A cast (HLMD Fr 1b) of a right mandible housed at the Hessisches Landesmuseum, Darmstadt, is erroneously labelled
“Simorre, holotype of Conohyus simorrensis” but the cast is of MNHN HGP 16 from Le Fousseret. This cast was studied by Hünermann (1968) and appears to have formed the basis for his understanding of the species. These two mandibular specimens from Le Fousseret
represent a single individual.
Measurements of fossils were made with sliding calipers to the nearest tenth of a mm. Images were taken with a Sony Cybershot
camera, and enhanced with Photoshop Elements 3.
The nomenclature of cusps, grooves and other structures in the teeth of suids follows the systems of Hünermann (1968), Pickford (1988) and Van der Made (1998). The meristic positions of teeth is given relative to the occlusal surface, represented by a forward slash (/). For upper
teeth the position is shown above the slash (eg M3/: upper third molar) and for lower teeth it is below the slash (eg p/4:
lower fourth premolar). Abbreviations of teeth are in upper case for upper teeth, and lower case for lower teeth: “I” and
“i” for incisors, “C” and “c” for canines, “P” and “p” for premolars, “M” and “m” for molars, and “D” and “d” for deciduous
cheek teeth.
It is noted that Stehlin (1899–1900) counted the premolars from back to front, thus his p/1 is what we would today call p/4,
and his p/2 is what we call p/3. Thus, his description of the cast of the mandible from Le Fousseret housed in the Bologna
Museum, as containing p/2-m/2 is confusing to those who do not realise the basis for his determination of meristic position.
In today’s parlance it contains only the p/3-m/2.
Bivariate and univariate plots in combined diagramsTOP
In the metric analyses provided in this paper, the breadth variation is given to the left of the bivariate plot (a thin vertical
line for each tooth position with the range of breadths represented by a thick line), and the length variation beneath the
bivariate plot (a horizontal line with the range of lengths represented by a thick line). This way of presenting the data
provides visual and metric access to the proportional differences that exist between different tooth positions, an important
source of information concerning tetraconodont teeth, because the inter-dental proportions help define the various taxa. Traditional
bivariate plots of each tooth position on its own fail to distinguish several of the taxa, which can have very similar molar
dimensions, for instance, but very different premolar dimensions. Likewise, traditional univariate plots fail to distinguish
some taxa, but when combined with bivariate plots the differences emerge. Whilst this new way of presenting the data appears
at first glance to be complicated, its advantages outweigh its inconveniences. The same measurement data are presented in
the bivariate plots (in which tooth positions are labelled) and in the univariate plots. It is stressed that the combination
employed in this paper yields a more coherent assessment of dental variation than either the bivariate or univariate plots
on their own. Once the reference bivariate plots have been prepared, fossils from other localities can be compared with them,
in this paper presented by letters of the alphabet in the univariate plots.
Institutional abbreviationsTOP
BSPG – Bayerische Staatsammlung für Paläontologie und Historische Geologie, München
DPZ – Department of Palaeontology, Zaragosa
GIU – Geology Institute, University of Belgrade
GPIT – Geologisches und Palaeontologisches Institut Tübingen
HLMD – Hessisches Landesmuseum, Darmstadt
IGGML – Institut für Geowissenschaften/Geologie de Montanuniversität, Leoben
IPS – Institut Català de Paleontoloia Miquel Crusafont, Sabadell
IPUW – Institut für Paläontologie, Universität Wien
LMJG – Landesmuseum Joanneum Graz
MHNT – Muséum d’Histoire Naturelle, Toulouse
MNHL – Museum of Natural History, Lisbon
MNHN – Muséum National d’Histoire Naturelle, Paris
MSIM – Museo San Isidro, Madrid
NHMB – Natural History Museum, Basel
NHMM – Naturhistorisches Museum, Mainz
NHMW – Naturhistorisches Museum, Wien
SMNS – Staatliches Museum für Naturkunde in Stuttgart
Biochronology of European tetraconodont localitiesTOP
Tetraconodont suids have been found at many localities in Europe (Figure 1, Table 1) ranging in age from MN 5 to MN 9. A list of the localities and their positions relative to the European Mammal Zonation
(Mein, 1986) is provided in Table 1.
|
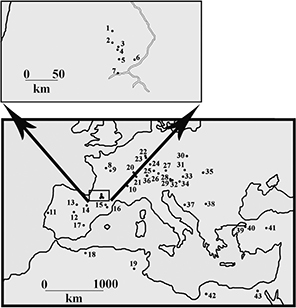 Fig. 1.— Location of selected European and North African tetraconodont localities. 1) Malartic, 2) Sansan, 3) Simorre, 4) Villefranche
d’Astarac, 5) Bonnefond, 6) Le Fousseret, 7) Saint Gaudens (Valentine), 8) Lublé, Hommes, 9) Channay, Pontlevoy, 10) La Grive-Saint-Alban,
11) Fonte do Pinheiro, Pero Filho, Azambujeira, 12) Carpetana, Puente de Vallecas, Alhambra, Somosaguas, 13) Montejo de la
Vega, 14) El Buste, La Ciesma, 15) St Quirze, 16) Can Llobateres, Can Ponsic, Santiga, 17) Mira, 18) Bou Hanifia (Dublineau),
19) Beglia, 20) Charmoille, 21) La Chaux-de-Fonds, 22) Gau-Weinheim (Wiesberg), 23) Esselborn, Eppelsheim, 24) Steinheim,
25) Breitenbrunn, 26) Urlau, Tutzing, 27) Pischelsberg, 28) Göriach, 29) Au, 30) Przeworno, 31) Klein Hadersdorf, Atzelsdorf,
32) Rosenthal 33) Neudorf, 34) Pitten, 35) Rudabanya, 36) Elgg, 37) Lucane, 38) Mala Miliva, 39) Nuri Yamut, 40) Pasalar,
41) Bâlâ, 42) Sahabi, 43) Wadi Natrun. Fig. 1.— Location of selected European and North African tetraconodont localities. 1) Malartic, 2) Sansan, 3) Simorre, 4) Villefranche
d’Astarac, 5) Bonnefond, 6) Le Fousseret, 7) Saint Gaudens (Valentine), 8) Lublé, Hommes, 9) Channay, Pontlevoy, 10) La Grive-Saint-Alban,
11) Fonte do Pinheiro, Pero Filho, Azambujeira, 12) Carpetana, Puente de Vallecas, Alhambra, Somosaguas, 13) Montejo de la
Vega, 14) El Buste, La Ciesma, 15) St Quirze, 16) Can Llobateres, Can Ponsic, Santiga, 17) Mira, 18) Bou Hanifia (Dublineau),
19) Beglia, 20) Charmoille, 21) La Chaux-de-Fonds, 22) Gau-Weinheim (Wiesberg), 23) Esselborn, Eppelsheim, 24) Steinheim,
25) Breitenbrunn, 26) Urlau, Tutzing, 27) Pischelsberg, 28) Göriach, 29) Au, 30) Przeworno, 31) Klein Hadersdorf, Atzelsdorf,
32) Rosenthal 33) Neudorf, 34) Pitten, 35) Rudabanya, 36) Elgg, 37) Lucane, 38) Mala Miliva, 39) Nuri Yamut, 40) Pasalar,
41) Bâlâ, 42) Sahabi, 43) Wadi Natrun.
|
|
Table 1—List of European Tetraconodont localities and their estimated correlations to the MN Zonation
| Locality |
Correlation this paper |
Locality |
Correlation this paper |
| Alhambra |
MN 5-6 |
Lucane |
MN 5 |
| Anwil |
MN 7/8 |
Mala Miliva |
MN 5 |
| Atzelsdorf |
MN 9 |
Malartic |
MN 7/8 |
| Au |
MN 5-6 |
Mira |
MN 9 |
| Azambujeira |
MN 8-9 |
Montejo de la Vega |
MN 5 |
| Bonnefond |
MN 8-9 |
Neudorf |
MN 6 |
| Breitenbrunn |
MN 7/8 |
Pasalar |
MN 6 (or 5) |
| Can Llobateres |
MN 9 |
Pero Filho |
MN 6 |
| Can Ponsic |
MN 9 |
Pischelsberg |
MN 6 |
| Channay |
MN 5 |
Pitten |
MN 6 |
| Charmoille |
MN 9 |
Pontlevoy |
MN 5 |
| El Buste |
MN 7/8 or 9 |
Przeworno |
MN 6 or 7/8 |
| Elgg |
MN 5 or 6 or 7 |
Puente de Vallecas |
MN 5 |
| Eppelsheim |
MN 8 + 9 + 10 |
Rosenthal |
MN 5-6 |
| Esselborn |
MN 9 |
Rudabanya |
MN 9 |
| Fonte do Pinheiro |
MN 9 |
Sansan |
MN 6 niveau répère |
| Gau Weinheim |
MN 8 + 9 |
Santiga |
MN 9 |
| Göriach |
MN 6 |
Simorre |
MN 7/8 |
| Hommes |
MN 5 |
Somosaguas |
MN 5 |
| Klein Hadersdorf |
MN 7 |
Steinheim |
MN 7 |
| La Chaux-de-Fonds |
MN 7/8 |
St-Gaudens (Valentine) |
MN 7/8 |
| La Ciesma |
MN 9 |
St Quirze |
MN 7/8 |
| La Grive-Saint-Alban |
MN 7/8 |
Tutzing |
MN 6 |
| Le Fousseret |
MN 7/8 |
Urlau |
MN 7/8 |
| Lublé |
MN 5 |
Villefranche d’Astarac |
MN 7/8 |
Systematic DescriptionsTOP
Family Suidae Gray, 1821
Subfamily Tetraconodontinae Lydekker, 1876
Genus Conohyus Pilgrim, 1925
Type species Sus simorrensis Lartet, 1851
Other species included:
Conohyus doati (Lartet, 1851)
Emended diagnosis of the genus:
Tetraconodont suids in which the anterior premolars (p/1, p/2, P1/, P2/) are not vertically offset in the jaw with respect
to the p/3 and P3/, crowns of anterior premolars slightly sectorial without marked mesial and distal buccal expansion and
with quite tall, pointed main cusp, anterior premolars markedly smaller than P3/ and P4/; occlusal surface of talonid of m/3
slightly twisted with respect to the surface of the anterior lophids, distal root of m/3 slightly inclined buccally, talonid
cusp positioned slightly buccal of the midline of the tooth; M3/ with posterior accessory cusplet slightly to the lingual
side of the midline of the crown, largely posterior to the hypocone; dentine-enamel junction of the molars have relatively
low relief (dentine exposure occurs with medium wear); lower male canines scrofic in section with shallow lingual and buccal
longitudinal gutters and a prominent longitudinal ridge of cementum on the enamel-free distal surface.
Differential diagnosis:
Conohyus differs from Retroporcus by the more sectorial anterior premolars set at the same level as the posterior premolars; by the presence of a longitudinal
ridge of cementum on the posterior surface of the lower canine; by its less symmetrical m/3 talonid and the buccally flaring
root of m/3. Conohyus differs from Parachleuastochoerus and Lophochoerus by its more inflated posterior premolars and less symmetrical m/3 crown. Conohyus differs from Sivachoerus by its more inflated posterior premolars with weak to absent accessory cusplets on the lingual side of the paracone and metacone,
and by the less vertical buccal surface of the P4/. Conohyus differs from Tetraconodon by its less inflated posterior premolars. Conohyus differs from Nyanzachoerus and Gerontochoerus by its more inflated posterior premolars and by the absence of elongation of the m/3 by addition of extra lophids.
Occurrence of the genus:
Austria (Ilz, St Oswald): France (Bonnefond, Castelnau d’Arbieu, Channay (Faluns), Doué-la-Fontaine, Gers, Hommes, La Grive-St-Alban,
Le Fousseret, Lublé, Meigné-le-Compte, Noyant-le-Plain, Pontigné, Pontlevoy, Simorre): Germany (Gunzberg, Hammerschmiede,
Hinterauerbach bei Wartenburg, Kleineisenbach, Neufeld, Obere Susswassermolasse, Pischelsberg, Tutzing, Urlau, Wartenberg
bei Erding): Poland (Przeworno): Portugal (Fonte de Pinheiro): Spain (Alhambra, Carpetana, El Buste): Switzerland (Braunkohle
Ostschweiz, Elgg, Hohrain-Waldi, Schlatt am Kohlfirst).
Species Conohyus simorrensis (Lartet, 1851)TOP
Synonymy:
partim Sus simorrensis Lartet, 1851; partim Sus (?) Doati Lartet, 1851; partim Hyotherium simorrense (Lartet, 1851): Stehlin, 1899–1900.
Diagnosis of the species:
Species of Conohyus in which the molar row measures ca 60 mm and the p/2-p/4 row measures ca 56 mm. Anterior premolars somewhat trenchant, distal
root of m/3 slightly buccally tilted. Lower male canine possesses a ridge of cementum on the distal enamel-free surface of
the crown.
Type material:
Lectotype: Stehlin’s concept (1899–1900) of Conohyus was primarily based on the left mandible from Le Fousseret, so, in order to respect the basis of his work and to promote
stability of nomenclature, we select as lectotype of Conohyus simorrensis, the mandible from Villefranche d’Astarac, which most closely resembles that from Le Fousseret. It consists of left and right
mandibles, the right one (MHNT PAL 2011.0.84.1) composed of two fragments, one containing p/2-p/3 (84.1.2) and the other the
p/4-m/2 (84.1.1). MHNT PAL 2011.0.84.2, a left mandible containing p/4-m/3 is the antimere of the right one and thus represents
the same individual.
Type horizon and type locality:
MN 6, Villefranche d’Astarac, France.
Occurrence of the species:
Austria (Ilz, St Oswald): France (Castelnau d’Arbieu, Channay (Faluns), Doué-la-Fontaine, Gers, La Grive-St-Alban, Le Fousseret,
Lublé, Pontlevoy, Simorre): Germany (Hammerschmiede, Hinterauerbach bei Wartenburg, Kleinei-senbach, Neufeld, Pischelsberg,
Urlau, Wartenberg bei Erding): Poland (Przeworno): Spain (Alhambra, Carpetana): Switzerland (Braunkohle Ostschweiz, Elgg).
Referred material:
Simorre: MNHN SML 76, left P3/; SML 37, right P4/, SML 57, left M2/; SML 36, left M3/, SML 72, right lower canine; SML 81, left D3/;
SML 80, p/3; SML 39, left p/4; SML 74, left m/3; SML 464, right P4/, SML 36, left M3/.
Le Fousseret: MHNT PAL 2012.0.206, left mandible containing p/3-m/2; MNHN HGP 16, right mandible with p/4-m/3.
Descriptions:
Lectotype: The most complete specimen from Villefranche d’Astarac comprises a right mandible in two parts stuck together with plaster-of-paris,
the anterior part containing p/2-p/3 (MHNT PAL 2011.0.84.1.2) and the posterior fragment containing p/4-m/2 (MHNT PAL 2011.0.84.1.1).
It is assumed that the anterior part of the jaw represents the same individual as the rear part, even though the contact between
the pieces is obscured under plaster, and despite the observation that the front section has greenish silty matrix and the
rear part a brownish silty matrix. Being the most diagnostic, the part containing the p/2 and p/3 is nominated lectotype.
The left mandible (MHNT PAL 2011.0.84.2) contains heavily worn p/4-m/3, and judging from the preservation characters and the
similar stage of wear of the teeth, it represents the same individual as the lectotype. The two mandibles can be described
together.
The jaw fragment, MHNT PAL 2011.0.84.1.2, contains p/2 and p/3. The p/2 is a slender, sectorial tooth considerably smaller
than the p/3. It has thin enamel, visible distally where flakes of enamel have broken off the tooth. The apex of the tooth
is unworn and pointed, the mesial accessory cusplet low and exposing dentine, there is no cingulum, and the enamel is wrinkled.
The cervix of the tooth is at the same level as that of the p/3 (ie there is no vertical offset between these two teeth, an
important point considering the other specimens from the locality). The roots are almost vertical and not particularly enlarged.
The p/3 is a large tooth, heavily worn, with thin enamel. The mesial accessory cusplet is lightly worn and has cingular extensions
buccally and lingually. The precristid is sharp and prominent, and the enamel wrinkled. The posterior accessory cusplet is
low, small and with cingular folds buccally and lingually.
The p/4 is heavily worn with coarsely wrinkled enamel, a low mesial accessory cusplet from which rounded cingula depart buccally
and lingually. The posterior accessory cusplet is worn, but was low and had cingular folds buccally and lingually. On the
buccal side there is a cingular swelling, but not forming a true cingulum, and on the lingual side there is no sign of a cingulum.
The anterior root is strongly developed.
The m/1 is very deeply worn. It has a large basal pillar in the buccal end of the median transverse valley.
The m/2 is heavily worn, showing four main cusps and well developed anterior, median and posterior accessory cusplets in the
midline of the tooth. There is a large but low basal pillar in the buccal end of the median transverse valley. The beaded
anterior cingulum is mesio-distally broad but fades out on the buccal and lingual surfaces of the crown. The enamel is wrinkled.
The m/3 is in heavy to medium wear with dentine exposed on the mesial cusp pair. The median accessory cusplet is compressed
mesio-distally but broad bucco-lingually. There is a clear buccal cingulum. The talonid complex is positioned slightly buccally
to the midline of the crown. The talonid is slightly twisted with respect to the two anterior lophids and this is reflected
in the orientation of the distal root, which leans slightly laterally. This is important to note because in other specimens
from Villefranche d’Astarac, the talonid of the m/3 is not twisted and the distal root is more vertically oriented in the
jaw.
Other material:
MNHN SML 76 is an unworn germ of a left P3/, which shows an apex that is subdivided into two points, a weak disto-lingual
shelf bordered by cingular margin, a well developed mesial cusplet flanked either side by short cingular folds, and the enamel
is wrinkled. Light wear would eradicate the evidence for the bifid apex.
The P4/, MNHN SML 37 possesses its roots, of which there are three, a large circular lingual one, and two subequal slightly
compressed buccal ones.
The only M1/s in the collection are fragmentary and worn.
M2/, MNHN SML 57 has four main cusps at the corners of the tooth and prominent mesial, median and distal accessory cusplets
in the midline. The mesial and distal cingula are beaded and do not reach the lingual and buccal surfaces of the crown. Both
ends of the median transverse valley show low basal pillars which form shallow foveae between them and the bigger median accessory
cusplet. There is quite marked buccal and lingual crown flare.
M3/, SML 36 shows a well developed buccal cingulum, above all on the buccal side of the paracone fading out distally. The
crown narrows distally due to the small structures around the hypocone and the better developed hypoconule (talon complex).
The four main cusps recall the same parts of the M2/ except that the second loph is narrower than the first one. Both specimens
have a small pointed basal pillar in the buccal end of the median transverse valley, and a bigger, flatter pillar in the lingual
end of the valley.
MNHN SML 72 is a right lower canine with a scrofic section. The lingual surface is much broader than the buccal surface. The
distal surface is devoid of enamel and neither does it have a ridge of cementum.
|
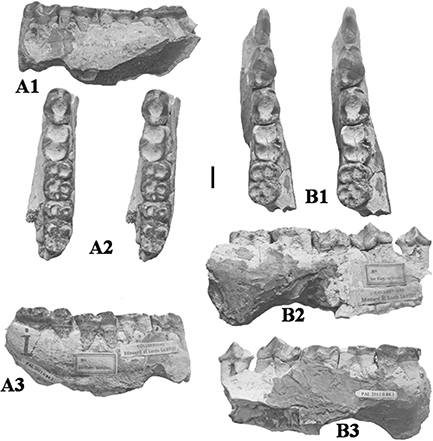 Fig. 2.— Conohyus simorrensis (Lartet, 1851) from Villefranche d’Astarac, France. A) MHNT PAL 2011.0.84.2 left mandible containing p/4-m/3, and B) MHNT PAL 2011.0.84.1,
right mandible composed of two fragments, one containing right p/2-p/3 (here nominated lectotype) (84.1.2) and the other the
p/4-m/2 (84.1.1). The left mandible is the antimere of the right one and thus represents the same individual. A1) buccal,
A2) stereo occlusal, A3) lingual views, B1) stereo occlusal, B2) buccal and B3) lingual views (scale: 10 mm). Fig. 2.— Conohyus simorrensis (Lartet, 1851) from Villefranche d’Astarac, France. A) MHNT PAL 2011.0.84.2 left mandible containing p/4-m/3, and B) MHNT PAL 2011.0.84.1,
right mandible composed of two fragments, one containing right p/2-p/3 (here nominated lectotype) (84.1.2) and the other the
p/4-m/2 (84.1.1). The left mandible is the antimere of the right one and thus represents the same individual. A1) buccal,
A2) stereo occlusal, A3) lingual views, B1) stereo occlusal, B2) buccal and B3) lingual views (scale: 10 mm).
|
|
MNHN SML 81 is interpreted to be left d/3 because of its tall triangular profile in lateral view. It has a small disto-lingual
swelling which has almost been eradicated by wear. The precristid and mesial accessory cusplet are also heavily affected by
wear. There are the bases of two roots.
|
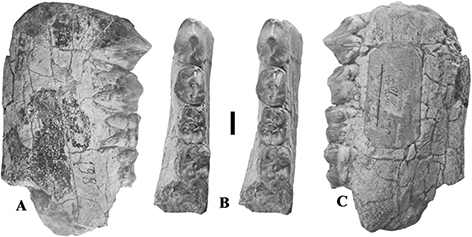 Fig. 3.— Conohyus simorrensis mandible from Le Fousseret, France. MHNT PAL.2012.0.206, (Leymerie 198 (Abadie, Fousseret)), left mandible containing p/3-m/2,
A) buccal, B) stereo occlusal and C) lingual views (scale: 10 mm). Fig. 3.— Conohyus simorrensis mandible from Le Fousseret, France. MHNT PAL.2012.0.206, (Leymerie 198 (Abadie, Fousseret)), left mandible containing p/3-m/2,
A) buccal, B) stereo occlusal and C) lingual views (scale: 10 mm).
|
|
MNHN SML 80 is a better preserved p/3 from Simorre, but it is broken lingually and mesially. The enamel is wrinkled, the main
cusp is tall (27.2 mm) with prominent pre- and post-cristids, and a small development of a lingual cristid descending from
the apex, but soon fading into the surface of the tooth. The distal cusplet is tall, and shows a lingual fold.
MNHN SML 39 is a left p/4 in medium wear. The mesial accessory cusplet is strong, with cingular folds either side of it, but
which fade out on the lateral and lingual sides of the tooth. The distal accessory cusplet is strong and taller than the mesial
one, and has broader folds either side, with depressions buccally and lingually separating it from the main cusp. The enamel
is wrinkled and the tooth has three roots, a strong one mesially, and two distally.
MNHN SML 74 is a left m/3 in light wear lacking its roots. It shows a well formed beaded cingulum on the buccal surface of
the protoconid which fades out distally before reaching the hypoconid. The talonid complex is positioned slightly to the buccal
side of the midline of the crown. In other respects the tooth is similar to the specimen from Malartic described below.
The following description is based on the Toulouse (MHNT PAL 2012.0.206) and Paris mandibles (MNHN HGP 16) from Le Fousseret
which represent a single individual. The wear pattern on the teeth is very close in the two specimens, and each of them has
a green label stuck to it with the mention “Chaerotherium (crossed out) N° 10, mach. inf. Fousseret Abadie”, and in different hand-writing the words “Sus simorrensis Lartet”. The colour of the teeth is the same, with the m/1s being considerably paler than the other teeth which are dark
grey apically becoming paler and browner cervically. The two specimens have similar matrix and in both specimens the mandible
has been extensively cracked, with sediment expansion damage, but not serious displacement of the fragments. Detailed descriptions
are provided because for Stehlin (1899–1900) the left mandible formed the basis of his concept of Hyotherium simorrense.
The left and right mandibles from Le Fousseret are poorly preserved and lack most of their bases. In the left mandible a small
section of the ventral margin is preserved beneath the m/2, and here the jaw is 55 mm deep measured from the gingival level.
The breadth of the jaw measured at the same place is 26.5 mm. The ascending ramus sweeps upwards starting at the level of
the front loph of m/3, and thus in lateral view hides the rearmost part of the m/3. There is a mental foramen beneath the
front of the p/4 at about half the height of the jaw.
The p/3 is lightly worn, with dentine showing down the postcristid of the main cusp, which is tall and located above the gap
between the roots. There is a marked pre-cristid which ends basally in a prominent accessory cusplet flanked by cingular folds.
Distally there is a small distal tubercle supported by tall cingular folds buccally and lingually, such that it is about half
the total height of the tooth. The enamel is wrinkled, and there is no sign of an interstitial contact facet caused by abrasion
with the p/2.
The p/4 is a robust tooth supported by two stout roots. It is moderately worn, exposing dentine from the apex of the main
cusp down the post-cristid and onto the distal accessory cusplet. The latter structure is flanked by broad cingular folds.
Anteriorly there is a precristid which terminates in a mesial accessory cusp and cingular folds. There is a prominent interstitial
contact facet caused by the p/3.
The m/1 is deeply worn, exposing much dentine on all four main cusps as well as the three accessory cusplets arranged mesially,
centrally and distally in the midline of the tooth. There is a well developed basal pillar in the buccal end of the median
transverse valley.
The m/2 is worn to the stage where small dentine lakes have formed on the four main cusps and the median accessory cusplet.
The mesial cingulum extends slightly onto the buccal surface where it shows three beads. The basal pillar in the buccal end
of the median transverse valley is likewise formed of three beads, now worn down. Distally, there are cingular cusplets either
side of the distal accessory cusplet.
The m/3 is constructed along the same lines as the m/2 except that behind the hypoconulid it possesses a stout talonid as
tall as the main cusps, with strong beaded cingular folds extending mesially onto the buccal and lingual sides. The talonid
is more or less in the midline of the crown. Because this tooth is less worn than the m/2, it shows remnants of the Fürchen
and the coarse enamel wrinkles that typify tetraconodont molars. The cingulum is well developed at the buccal end of the median
transverse valley, and on the mesial half of the protoconid.
Synonymy:
Sus (?) Doati Lartet, 1851; Hyotherium simorrense Var. Doati (Lartet, 1851): Stehlin, 1899–1900; Hyotherium simorrense var. doati (Lartet, 1851): Roman, 1907; Conohyus ebroensis Azanza, 1986.
Non Conohyus giganteus (Falconer & Cautley, 1847): Van der Made 1998.
Diagnosis of the species:
Large species of Conohyus; m/1-m/3 ca 68 mm.
Type material:
Lectotype HLMD cast Fr3, right M3/. (The current whereabouts of the original fossil are unknown).
Type horizon and type locality:
MN 8–9, Bonnefond, Haute Garonne, France.
Occurrence of the species:
France (Bonnefond); Germany (Gau-Weinheim; Obere Süsswassermolasse (Wartenberg bei Erding)); Portugal (Fonte de Pinheiro);
Spain (El Buste, ?La Ciesma).
Referred material:
Bonnefond: MNHN HGP 66, right lower canine; Lost left m/3 figured by Stehlin (1899–1900, pl. 1, fig. 9).
Fonte de Pinheiro: MNHL, left mandible with p/2-m/3, rt p/3, m/2 and m/3 (cast in NHMB SPA).
El Buste: DPZ 2001, right mandible with p/4-m/2.
Wartenberg bei Erding (Obere Süsswassermolasse): BSPG 1952.I.279, left M3/
Dinotheriensande: HLMD Din 89, left M3/; NHMM 1932/402, right m/3; NHMM 1932/490, left M3/; NHMM 1952/582, right M3/
RemarksTOP
Large specimens of Conohyus reported from France and Spain (Conohyus doati, Conohyus ebroensis and some of the material previously identified as Conohyus simorrensis) belongs to Conohyus doati (Lartet, 1851). The type locality of this species is Bonnefond (Lartet, 1851). The greater dimensions of its teeth compared to those of Conohyus simorrensis, an observation which formed part of the “diagnosis” of the species by Lartet (1851) is borne out by subsequent discoveries in Spain (Roman, 1907; Golpe-Posse, 1972; Azanza, 1986). Descriptions of the main fossils are given below.
BonnefondTOP
The molars of Conohyus from Bonnefond (Stehlin, 1899–1900) are larger than the Conohyus specimens from Villefranche d’Astarac and Simorre, a fact already noted by Lartet (1851) who, on this basis, defined the species Sus (?) doati. Stehlin (1899–1900) illustrated two specimens as “types” of the species, a right M3/ (Stehlin, 1899–1900, pl. 1, fig. 7) and a left m/3 (Stehlin, 1899–1900, pl. 1, fig. 9). The upper third molar is here nominated the lectotype. The lower canine from Bonnefond (MNHN HGP 66) is larger than that
from Carpetana, Spain (Pickford, 2013) but like it, it possesses a band of cementum on the distal enamel-free surface of the crown, a unique feature among suids.
|
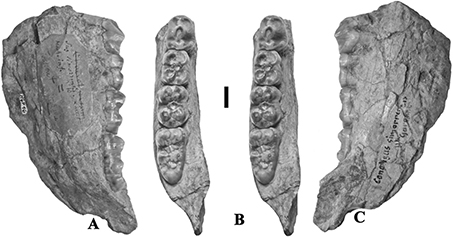 Fig. 4.— Conohyus simorrensis jaw from Le Fousseret, France. MNHN HGP 16 (Leymerie N° 10, Abadie, Fousseret) right mandible containing p/4-m/3, A) lingual,
B) stereo occlusal and C) buccal views (scale: 10 mm). Fig. 4.— Conohyus simorrensis jaw from Le Fousseret, France. MNHN HGP 16 (Leymerie N° 10, Abadie, Fousseret) right mandible containing p/4-m/3, A) lingual,
B) stereo occlusal and C) buccal views (scale: 10 mm).
|
|
|
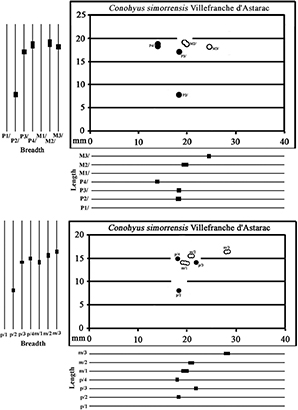 Fig. 5.— Bivariate and univariate plots of the upper teeth (top) and lower teeth (bottom) of Conohyus simorrensis from the type locality, Villefranche d’Astarac, France (premolars: solid symbols; molars: hollow symbols). Fig. 5.— Bivariate and univariate plots of the upper teeth (top) and lower teeth (bottom) of Conohyus simorrensis from the type locality, Villefranche d’Astarac, France (premolars: solid symbols; molars: hollow symbols).
|
|
Fonte do PinheiroTOP
A suid jaw from the fluvial deposits of the Palaeo-Tagus at Fonte do Pinheiro, Portugal, was described by Roman (1907). The specimen comprises a reasonably complete left mandible containing the p/2 (damaged), p/3-m/3 and isolated right p/3,
m/2-m/3. It was attributed to Hyotherium simorrense var. doati on account of its large dimensions, comparable to the specimens from Bonnefond, France, the type locality of the species
Sus doati (Lartet, 1851), discussed by Stehlin (1899–1900).
Van der Made (1989) included the Fonte do Pinheiro specimen in Conohyus ebroensis Azanza (1986) because of the similarities in dimensions with material from the type locality of this species, El Buste, Spain.
El BusteTOP
The question concerning the validity of the species Conohyus ebroensis arises. It is close in dimensions to four named species
of tetraconodont – Genus indet. antediluvianus (Kaup, 1833) from the Dinotheriensande, Conohyus doati (Lartet, 1851) from Bonnefond, Conohyus melendezi Golpe-Posse (1972) from Mira, and to “Sus” valentini Filhol 1882, from St-Gaudens (the two latter taxa are here considered to belong to the
genus Parachleuastochoerus as the new combination Parachleuastochoerus valentini (Filhol, 1882)). Conohyus ebroensis is compatible in dimensions to the Bonnefond material and to the lower jaw from Fonte do Pinheiro and
the species is here considered to be a junior synonym of Conohyus doati.
The mandible from El Buste possesses a deep sublingual fossa beneath the molar row, shallowing beneath the p/4 before joining
the genial fossa. The symphysis, which is not fused in this sub-adult individual (p/4 in the process of erupting) extends
back as far as the rear of p/2. The front of the jaw is broken, but shows parts of the alveoli for the central and second
incisors, the small root (probably a female individual) and a two-rooted p/1. There is a short diastema between the p/1 and
the p/2. There are two mental foramina, one beneath the front root of p/2, the other in the upper third of the jaw beneath
the p/4.
Conohyus doati is still poorly known. It is close morphologically to Conohyus simorrensis but is substantially larger, as originally observed by Lartet (1851). The lower male canine is scrofic in section and sports an elongated ridge of cementum on the enamel-free distal surface
(Stehlin, 1899–1900), as does the canine of Conohyus simorrensis from Carpetana, Spain. Other tetraconodonts with scrofic lower canines show no such ridge of cementum.
It should be noted that the lectotype upper third molar from Bonnefond (27,5×20 mm) is considerably smaller than the M3/ in
the type specimen of Sivachoerus giganteus (Falconer & Cautley, 1847) (which was identified as Conohyus giganteus by Van der Made, 1998, 2004) from the Siwaliks of the Indian Subcontinent (M3/, 42.3×29 mm) which makes it unlikely that the large European Conohyus specimens belong to the Indian species (Van der Made, 1998, 2004).
Golpe-Posse (1972) erected two new species of Conohyus, 1) C. melendezi, on the basis of fossils from Mira, Spain, which is large (Van der Made, 1998) and belongs to Parachleuastochoerus valentini (new combination) and 2) C. cuspidatus from Armantes I and Munebrega AB (Spain) which are small and belong to Hyotherium soemmeringi (Van der Made, 1990).
Azanza (1986) erected the species Conohyus ebroensis for specimens which are somewhat larger than Conohyus simorrensis. Van der Made (1998) attributed the specimens to Conohyus giganteus (Falconer & Cautley, 1847) but the latter species is considerably larger than any of the Conohyus specimens from Europe.
It is concluded that the tetraconodont mandibles from Fonte do Pinheiro and El Buste belong to Conohyus doati (Lartet, 1851).
Conohyus sp.TOP
Isolated or damaged teeth that probably belong to Conohyus simorrensis:
Simorre: MNHN Si 322, right I3/; MNHN SML 34, right upper canine; SML 75, left M2/, SML 79, left M2/; SML 323, left lower
canine; SML 269, right m/1; SML 73, left m/2; SML 1144, left m/2; SML 77, right m/2.
Description:
Some isolated or heavily worn tetraconodont teeth from Simorre and Rajegats cannot be attributed with certainty to a species.
They are identified as Conohyus sp.
An isolated upper incisor, MNHN Si 322, is interpreted to be a right I3/ It has a broad distal crest descending from the apex,
and a sharp lingual crest leading distally that closes off an elongated distal fovea. The protocrista ends in a low fold directed
lingually.
The upper canine MNHN SML 34 is bucco-lingually compressed, is triangular in buccal view and has a root with a prominent lingual
groove. There is a cementum deposit buccally. The posterior crista is sharp, and flanked by a zone on the buccal surface of
the tooth which is devoid of enamel. The protocrista is sharp at its base but has been worn away towards the apex by contact
with the lower canine.
The two M2/s from Simorre (MNHN SML 75, 79) vary slightly in dimensions and wear, but are morphologically similar to each
other.
MNHN SML 323 is a deeply worn left central lower incisor, showing a prominent lingual crest and strong mesio-distal compression.
The only m/1s in the sample are broken, MNHN SML 269 being the front half of an unworn tooth showing the post-cristid of the
metaconid detached apically to form a separate small cusplet, and Fürchen 2 and 3 meeting to carve off a small accessory cusp
between the metaconid and the median accessory cusplet.
Three second lower molars in the collection (MNHN SML 73, 1144, 77) can be described together. SML 73 is unworn, shows wrinkled
enamel, the detached post-cristid of the metaconid, and the accessory cusp between the metaconid and the median accessory
cusplet. The mesial cingulum extends onto the buccal surface where it forms a series of beads before fading out distally.
There is a cingular remnant in the buccal end of the median transverse valley but the lingual end of the valley is open. The
hypoconulid is robust and flanked by two low cingular crests that lead mesio-lingually and mesio-buccally to close off two
small foveae.
Genus Retroporcus nov.TOP
Type species:
Retroporcus complutensis nov. sp.; Retroporcus sindiensis (Lydekker, 1884); Hyotherium soemmeringi matritensis nov. sp. Golpe-Posse, 1972, pp. 122, 155–156, pl. 6, fig. 6b.
|
 Fig. 6.— HLMD Fr 3, cast of the lectotype right M3/ of Conohyus doati from Bonnefond, France, occlusal view (scale: 10 mm). Fig. 6.— HLMD Fr 3, cast of the lectotype right M3/ of Conohyus doati from Bonnefond, France, occlusal view (scale: 10 mm).
|
|
Occurrence of the genus:
Austria (Au bei Aufkenz, Göriach): France (Auverse, Doué-la-Fontaine, Faluns, Lasse, Malartic, Simorre (Rajegats), Villefranche
d’Astarac): Germany (Eppelsheim, Gau Weinheim, Laichingen): India (Siwaliks); Pakistan (Siwaliks): Serbia (Mala Miliva): Spain
(Puente de Vallecas, Somosaguas): Switzerland (Anwil): Turkey (Pasalar)
Derivatio nominis:
Retro from the Latin in the sense of “going backward in time”, porcus, Latin for pig.
Species Retroporcus complutensis nov.TOP
Synonymy:
Conohyus simorrensis simorrensis (Lartet, 1851): Petronijevic, 1967; Conohyus simorrensis goeriachensis Van der Made 1989: Van der Made & Salesa, 2004
Diagnosis:
Species of Retroporcus slightly smaller than Retroporcus matritensis, with shorter premolars, in particular p/1, m/1-m/3 about 55 mm and p/2-p/4 about 50 mm.
Type material:
MNCN SOM N Cata 2 2008–83, left mandible containing p/2-m/3.
Type horizon and type locality:
MN 5, Somosaguas, Madrid, Spain.
Occurrence of the species:
Spain (Somosaguas), Serbia (Mala Miliva)
Referred material:
Somosaguas: MNCN SOM N Cata 2 2008–83, left mandible with p/2-m/3; Cata 2 10 101, right i/1; N 2002 203, right i/1; Cata 2 07.6, right
i/2; N 2002 1030, left p/1; S 2005 3, left m/1; N 12 414–87, left P3/; MNCN SMCN 3, left m/3; N 2002 1079, left P1/; N 98
00 324, left P3/; N 2002 1109, left M1/; S 1998 112, right M1/; N 2001 484, left M2/; N 2001 976, right m/2; N 1998 83, right
M3/.
Mala Miliva: GIU Belgrade, left mandible with p/1-m/2, right mandible with p/1-m/3.
Derivatio nominis:
The species name is for the Universidad Complutense, Madrid, in the grounds of which is situated Somosaguas where the type
material was excavated.
Descriptions:
Somosaguas
Holotype:
SOM CATA 2’08–83 is an adult left mandible containing p/2 to m/3 in light wear. The jaw is deep and rather narrow (48,5 mm
deep beneath m/1, 21 mm broad at m/1) with a deeply excavated sub-lingual fossa that extends anteriorly almost to the rear
of the symphysis. The genial fossa is at the level of the back of the p/2. There are two mental foramina, one beneath the
gap between p/2-p/3, the other beneath the rear of p/3, both are about mid-height of the jaw. There is a superior transverse
torus slightly overhanging the genial fossa.
The p/2 has two roots that are parallel to each other and steeply inserted in the jaw. The crown has a long posterior projection
of the crown which is slightly lower than the cervix of the p/3. The main cusp is positioned slightly mesial to the gap between
the two roots, making the pre-cristid shorter than the post-cristid. There is a well formed mesial accessory cusplet with
light cingular folds buccally and lingually. The post-cristid bifurcates about half way from the apex of the main cusp towards
the distal margin of the crown, sending a low ridge linguo-distally, which makes the rear of the tooth broader than the front.
The disto-buccal surface of the tooth is slightly swollen.
The p/3 is much larger than the p/2, its mesial accessory cusplet being taller than the entire p/2 in front of it and the
main cusp about twice the height of the p/2. The crown is posed on two stout roots that slant slightly anteriorly. The main
cusp is located above the rear root, which makes the pre-cristid longer than the post-cristid. The mesial accessory cusp is
about half the height of the tooth, and has cingular folds lingually and buccally that border off low depressions behind them.
The posterior accessory cusplet is taller than the mesial one, and is more voluminous with thick cingular folds extending
buccally and lingually. The buccal enamel is heavily wrinkled and the enamel extends further rootwards buccally than lingually,
especially on the distal root.
The p/4 is shorter than the p/3 but has approximately the same distal breadth. Its main cusp is also slightly lower than that
of the p/3. The mesial accessory cusplet is about half the height of the tooth. The buccal cingular fold is strong and vertically
oriented, but the lingual one is weak. The distal accessory cusplet is better individualised in the p/4 than in the p/3, and
its cingular folds emerge from its rear edge. Narrow but quite deep grooves separate the distal accessory cusplet from the
main cusp, especially marked on the buccal side of the crown. The enamel extends further rootwards buccally than lingually
especially on the distal root.
The m/1 is damaged by cracking and displacement of the lingual half relative to the buccal half.
The m/2 is also cracked but the two parts are close together. The crown shows the usual four main cusps arranged in two lophids,
with well formed mesial, central and distal accessory cusplets disposed along the mid-line of the tooth. The distal accessory
cusp is large, the central one bucco-lingually broad, but mesiodistally compressed, and the mesial cusplet is small. The mesial
cingulum is sharp and beaded and there are tiny cingular remnants in the buccal end of the median transverse valley.
The m/3 is lightly worn. The two anterior lophids are close in size and morphology to the m/2, but there is a prominent talonid
complex comprised of the distal accessory cusplet accompanied distally by a large talonid cusp flanked by large cingular cusplets
lingually and buccally. The talonid cusp is slightly to the buccal side of the midline of the crown. The buccal cingulum is
well developed on the front half of the anterior lophid but fades out distally, but does leave a small remnant in the buccal
end of the median transverse valley. In distal view the distal root of the m/3 is observed to slant slightly towards the buccal
side of the jaw.
Other material:
MNCN Som-N-83 the isolated lower left third molar is heavily worn, but it is possible to observe that the talonid is located
slighty buccal to the midline of the tooth, and that its occlusal surface appears not to be twisted with respect to the surface
of the two anterior lophids.
The upper third molar from Somosaguas (Van der Made & Salesa, 2004) has a short talon in line with the lingual cusps of the two anterior lophs. The mesial cingulum is broad and wraps around
the protocone. The P3/ germ from the site shows a bifid apex which would disappear with a light degree of wear. The slightly
damaged left P2/ is bunodont with a low main cusp, separated from the posterior part of the tooth by a shallow cleft similar
to specimens of Retroporcus matritensis from Göriach (ex Conohyus simorrensis goeriachensis) and Villefranche d’Astarac.
RemarksTOP
As was already noted by Van der Made & Salesa (2004) the Somosaguas tetraconodont is smaller than Conohyus simorrensis goeriachensis (ie Retroporcus matritensis of this paper) but is morphologically similar to it.
Petronijevic (1967) described two relatively complete mandibular rami from Mala Miliva, Serbia, which he attributed to Conohyus simorensis simorrensis. The teeth are close in dimensions to the fossils from Somosaguas, Spain, with the exception of the p/1 which is shorter
in the Mala Miliva sample. The posterior premolars have the proportions of Retroporcus nov. gen. but overall the material from these two sites is smaller than that from Puente de Vallecas and Göriach. They are
here attributed to the new species, Retroporcus complutensis nov. gen. nov. sp.
Species Retroporcus matritensis nov. comb. (Golpe-Posse, 1972)TOP
Synonymy:
partim Hyotherium simorrense (Fraas, 1870): Stehlin, 1899–1900; Hyotherium soemmeringi matritensis nov. sp. Golpe-Posse, 1972; Conohyus simorrensis goeriachensis subnov. sp. Van der Made, 1989.
Diagnosis of the species:
Species of Retroporcus in which the lower molar row is ca 60 mm long, lower p/2-p/4 row measures ca 66 mm long; upper molar row measures ca 55 mm;
upper premolar row (P2/-P4/) measures ca 50 mm. Anterior premolars bunodont, vertically offset with respect to the level of
the p/3 and P3/. Distal root of m/3 not or only slightly buccally tilted. No signs of a ridge of cementum on the distal enamel-free
surface of the lower male canine.
Type material:
IPS 1868, left maxilla containing P4/-M3/ (Golpe-Posse, 1972, pl. 6, fig. 6b and Morales & Soria, 1985, pl. 3, fig. 4).
|
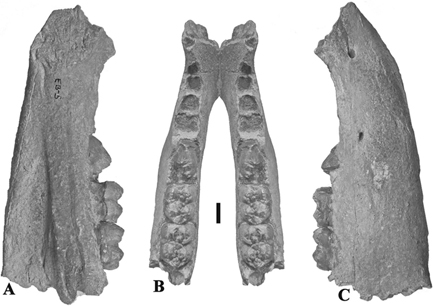 Fig. 7.— DPZ 2001, El Buste right mandible of Conohyus doati, A) lingual, B) occlusal view with mirror image aligned along the surface of the symphysis, C) buccal view (scale: 10 mm). Fig. 7.— DPZ 2001, El Buste right mandible of Conohyus doati, A) lingual, B) occlusal view with mirror image aligned along the surface of the symphysis, C) buccal view (scale: 10 mm).
|
|
Type horizon and type locality:
MN 5, Puente de Vallecas, Spain.
Occurrence of the species:
Austria (Au bei Aufkenz, Göriach): France (Auverse, Doué-la-Fontaine, Faluns, Lasse, Malartic, Simorre (Rajegats), Villefranche
d’Astarac): Germany (Eppelsheim, Gau Weinheim, Laichingen): Spain (Puente de Vallecas): Switzerland (Anwil): Turkey (Pasalar).
Referred material:
Puente de Vallecas: MSIM 2421, right mandible containing p/4-m/3; MSIM 2423, right m/3; MSIM 2680, left M1/; MSIM 2422, right M2/; MSIM 2424,
left M3/; MSIM 3286, right p/3; MSIM 2425, left p/4; MSIM 2421, right p/4; MSIM 2642, left P4/; MSIM 2680, left P4/.
Villefranche d’Astarac: MHNT PAL 2012.0.197, left mandible with p/2-p/3 and isolated p/4; PAL 2012.0.195, left m/1; PAL 2010.84.3, left m/3 in mandible
fragment; PAL 2012.0.196, right maxilla with P2/-P3/ and isolated P4/; PAL 2012.0.199, right D4/; PAL 2012.0.198, right M2/;
PAL 2012.0.194, right maxilla with P4, M2/ and isolated M3/.
Simorre: MNHN Si 319, left maxilla with P1/-P3/; Si 320, right P1/; Si 321, left P1/; MNHN SML 463, right P3/; SML 78, left P4/;
SML 234, left P4/; SML 337, right P4/; SML 324, right D4/; SML 290, right M3/.
Description:
Puente de Vallecas
Some of the fossils from Puente de Vallecas were briefly described by Golpe-Posse (1972) and Morales & Soria (1985). The left maxilla, IPS 63 (old number IPS 1868) contains the P4-M3. There are two distal alveoli for the P3/ but the tooth
is missing. The paracone and metacone of the P4/ are so close together, that even with the slight wear that the tooth has
suffered they have almost fused into a single cusp, as was noted by Golpe-Posse (1972). The anterior cingulum of the P4/ is well developed and ends buccally in a detached parastyle. The distal cingulum is strong
behind the protocone, but fades out buccally. There is a postmetacrista, almost forming a metastyle on the buccal surface
of the tooth. The first and second molars are damaged and deeply worn. The talon of the M3/ is short, and buccally there is
a complex of several small, low cusplets which impart a rounded rectangular outline to the occlusal outline. In the lingual
end of the median transverse valley there is a prominent accessory cusplet. The buccal cingulum is weak, but produces a small
cusplet in the buccal end of the median transverse valley, and a weak swelling on the buccal sides of the paracone and metacone.
The anterior and medial accessory cusplets are prominent, but the distal one is weakly developed and bordered distally by
a beaded cingulum.
MSIM 2421 is a right mandible with p/4-m/3. The p/4 is a large, inflated tooth with the Innenhugel so close to the protoconid
that with slight wear it has fused with it. The m/3 MSIM 290, has a centrally positioned talonid.
Villefranche d’Astarac
Lower dentition:
MHNT PAL 2012.0.197 is a left mandible fragment containing p/2-p/3 and an isolated p/4 which has interstitial wear facets
that indicate that it belongs to the same individual. The p/2 is lower crowned than the p/3 and it is is offset vertically
with respect to the p/3 such that its crown is aligned with the root of the p/3. This results in a distinct step in the occlusal
surface of the tooth row. The main cusp of the p/2 is positioned above the gap between the two roots, thus in the anterior
third of the tooth. The mesial accessory cusplet forms a low point from which low cingular folds depart buccally and lingually.
The postcristid is long and bifurcates distally, sending a low rounded crest buccally. There is a slight bend in the cervix
of the tooth above the gap between the two roots on the buccal and lingual sides. The roots are stout, the posterior one being
enlarged, the mesial one smaller and both slightly inclined mesially.
The p/3 is a taller tooth with stout roots and the main cusp in the middle of the tooth. The mesial accessory cusplet is low
and bordered by buccal and lingual cingular crests. The precristid is low and rounded due to light wear. The postcristid is
worn away, with enamel exposed as far as the posterior accessory cusplet. This cusplet is low and is bordered by low, rounded
cingular folds. In buccal and lingual views the cervix is observed to rise crownwards to a marked degree above the gap between
the two roots. The enamel extends further towards the root on the buccal side than it does on the lingual one. The enamel
is wrinkled. The two stout roots are inclined mesially.
|
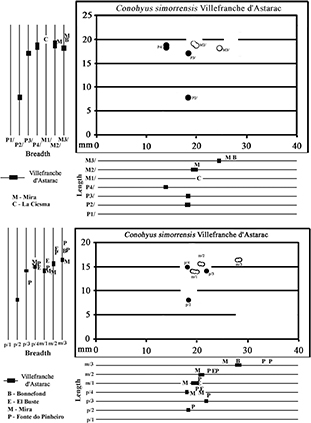 Fig. 8.— Metric comparison of the upper (top) and lower cheek teeth (bottom) of Conohyus simorrensis and Conohyus doati (circles – Villafranche d’Astarac; letters – other localities) (premolars – solid symbols; molars – hollow symbols). Fig. 8.— Metric comparison of the upper (top) and lower cheek teeth (bottom) of Conohyus simorrensis and Conohyus doati (circles – Villafranche d’Astarac; letters – other localities) (premolars – solid symbols; molars – hollow symbols).
|
|
The p/4 is heavily worn, and shows a prominent posterior accessory cusplet with buccal and cingular folds walling off lingual
and buccal depressions between the main cusp and the posterior accessory cusplet. The cervix is bent upwards above the gap
between the two roots. The enamel extends further rootwards on the buccal side than the lingual one. The roots are stout.
The left m/1 (MHNT PAL 2012.0.195) lacks the disto-lingual cusp (entoconid). The crown is in light to medium wear, and shows
four bunodont main cusps, and strongly developed median and posterior accessory cusplets. The anterior accessory cusplet is
weak, as is the mesial cingulum. There is a prominent basal pillar in the buccal end of the median transverse valley.
|
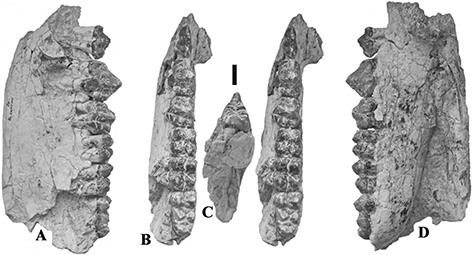 Fig. 9.— MNCN SOM N Cata 2 2008–83, holotype left mandible of Retroporcus complutensis, nov. gen. nov. sp. from Somosaguas, Madrid, Spain. A) buccal, B) stereo occlusal view, C) posterior view to show the slight
buccal tilt of the distal root in m/3, D) lingual view (scale: 10 mm). Fig. 9.— MNCN SOM N Cata 2 2008–83, holotype left mandible of Retroporcus complutensis, nov. gen. nov. sp. from Somosaguas, Madrid, Spain. A) buccal, B) stereo occlusal view, C) posterior view to show the slight
buccal tilt of the distal root in m/3, D) lingual view (scale: 10 mm).
|
|
MHNT PAL 2010.84.3 is a left m/3 in a fragment of mandible. The tooth is deeply worn exposing dentine in all the main cusps
and the median and posterior accessory cusplets. The wear pattern so produced shows almost circular dentine islands separated
from each other by thick enamel walls. There is a basal pillar in the buccal end of the median transverse valley. The talonid
consists of a low cusp positioned slightly to the buccal side of the midline of the crown, directly behind the hypoconulid
(posterior accessory cusplet) and it has well developed cingula running anteriorly on the buccal and lingual sides.
|
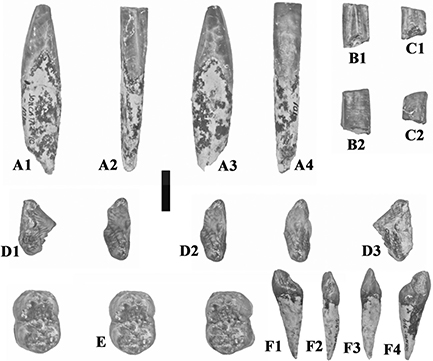 Fig. 10.— Isolated teeth of Retroporcus complutensis nov. gen. nov. sp. from the type locality, Somosaguas, Spain. A) MNCN SOM CATA 2’10 101, right i/1, A1 – mesial, A2 – lingual,
A3 – distal, A4 – labial views, B) MNCN SOM N 2002–2031, right i/1, B1 – lingual, B2 – labial views, C) MNCN SOM CATA 2’07
6, right i/2, C1 – lingual, C2 – labial views, D) MNCN SOM N 2002 1030, left p/1, D1 – buccal, D2 – stereo occlusal, D3 –
lingual views, E) MNCN SOM S 2005–3, left m/1, stereo occlusal view, F) MNCN SOM N 12 414–87, left I3/, F1 – lingual, F2,
distal, F3 – mesial, F4 – labial views. (scale: 10 mm). Fig. 10.— Isolated teeth of Retroporcus complutensis nov. gen. nov. sp. from the type locality, Somosaguas, Spain. A) MNCN SOM CATA 2’10 101, right i/1, A1 – mesial, A2 – lingual,
A3 – distal, A4 – labial views, B) MNCN SOM N 2002–2031, right i/1, B1 – lingual, B2 – labial views, C) MNCN SOM CATA 2’07
6, right i/2, C1 – lingual, C2 – labial views, D) MNCN SOM N 2002 1030, left p/1, D1 – buccal, D2 – stereo occlusal, D3 –
lingual views, E) MNCN SOM S 2005–3, left m/1, stereo occlusal view, F) MNCN SOM N 12 414–87, left I3/, F1 – lingual, F2,
distal, F3 – mesial, F4 – labial views. (scale: 10 mm).
|
|
Upper dentition:
The right maxilla in the Lartet collection (MHNT PAL 2012.0.196) contains the P2/ and P3/, and there is an isolated P4/ which
belongs to the same individual judging from the compatible interstitial wear facets of the teeth. The P2/ is a low crowned
tooth which is inserted in the maxilla well below the cervix of the P3/, which means that there is a prominent step or offset
in the occlusal surface of the tooth row. There is almost no bend in the cervix on the lingual side, but a slight bend is
present on the buccal side of the tooth. P2/ has two stout roots which are inclined slightly anteriorly, with the main cusp
positioned above the gap between the two roots. The crown is moderately worn, with dentine exposed apically and in the distal
cusplet complex. The mesial accessory cusplet is low, with cingular crests buccally and lingually. The distal accessory cusplet
is deeply worn as is the disto-buccal end of the postcrista.
|
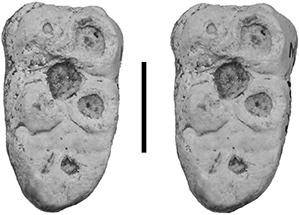 Fig. 11.— MNCN SMCN 3, stereo occlusal view of left m/3 of Retroporcus complutensis nov. gen. nov. sp. from Somosaguas, Madrid, Spain (scale: 10 mm). Fig. 11.— MNCN SMCN 3, stereo occlusal view of left m/3 of Retroporcus complutensis nov. gen. nov. sp. from Somosaguas, Madrid, Spain (scale: 10 mm).
|
|
The P3/ is a large tooth, triangular in occlusal outline. It has three roots, one stout one anteriorly, and two posterior
ones. The tooth is deeply worn, which has eradicated most of the morphology, with the anterior wear facet extending onto the
root of the tooth, but it is possible to make out that there was a mesial accessory cusplet bordered by buccal and lingual
cingular folds. Above the gap between the mesial and distal roots the cervix bends rootwards to a slight extent on both the
buccal and lingual sides of the tooth.
|
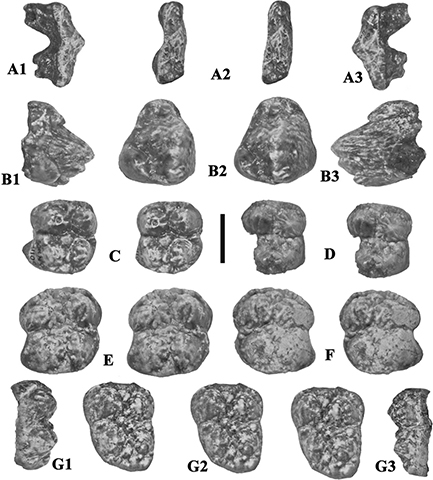 Fig. 12.— Isolated upper teeth of Retroporcus complutensis nov. gen. nov. sp. from the type locality, Somosaguas, Spain. A) MNCN SOM N 2002–1079, left P1/, A1 - lingual, A2 - stereo
occlusal, A3 - buccal views, B) MNCN SOM N 98-00-324, left P3/, B1 – lingual, B2 – stereo occlusal, B3 – buccal views, C)
MNCN SOM N 2002–1109, left M1/, stereo occlusal view, D) MNCN SOM S 1998–112, right M1/, stereo occlusal view, E) MNCN SOM
N 2001–484, left M2/ stereo occlusal view, F) MNCN SOM N 2001–976 right M2/, stereo occlusal view, G) MNCN SOM N 1998–83,
right M3/, G1 - buccal, G2 - stereo occlusal view, G3 – buccal views (scale: 10 mm). Fig. 12.— Isolated upper teeth of Retroporcus complutensis nov. gen. nov. sp. from the type locality, Somosaguas, Spain. A) MNCN SOM N 2002–1079, left P1/, A1 - lingual, A2 - stereo
occlusal, A3 - buccal views, B) MNCN SOM N 98-00-324, left P3/, B1 – lingual, B2 – stereo occlusal, B3 – buccal views, C)
MNCN SOM N 2002–1109, left M1/, stereo occlusal view, D) MNCN SOM S 1998–112, right M1/, stereo occlusal view, E) MNCN SOM
N 2001–484, left M2/ stereo occlusal view, F) MNCN SOM N 2001–976 right M2/, stereo occlusal view, G) MNCN SOM N 1998–83,
right M3/, G1 - buccal, G2 - stereo occlusal view, G3 – buccal views (scale: 10 mm).
|
|
The P4/ is heavily worn, but it is possible to deduce that it had two main cusps (one buccal, the other lingual) with pre-
and post-crista leading from the buccal cusp towards the midline of the tooth anteriorly and posteriorly respectively. There
are strong mesial and distal cingula.
|
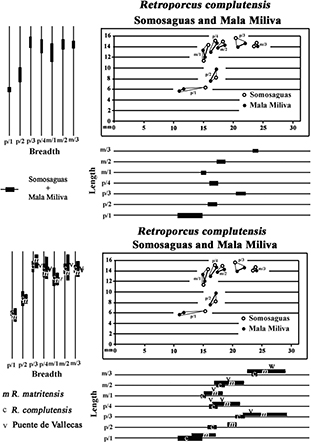 Fig. 13.— Bivariate and univariate plots of the lower cheek teeth of Retroporcus complutensis, nov. gen. nov. sp. from Somosaguas and Mala Miliva (premolars – solid symbols; molars – hollow symbols). Fig. 13.— Bivariate and univariate plots of the lower cheek teeth of Retroporcus complutensis, nov. gen. nov. sp. from Somosaguas and Mala Miliva (premolars – solid symbols; molars – hollow symbols).
|
|
The right D4/ (MHNT PAL 2012.0.199) has a slightly trapezoidal occlusal outline, with four main cusps and weak mesial, median
and distal accessory cusplets. The enamel is heavily wrinkled. Wear has exposed dentine at the apices of the four main cusps.
|
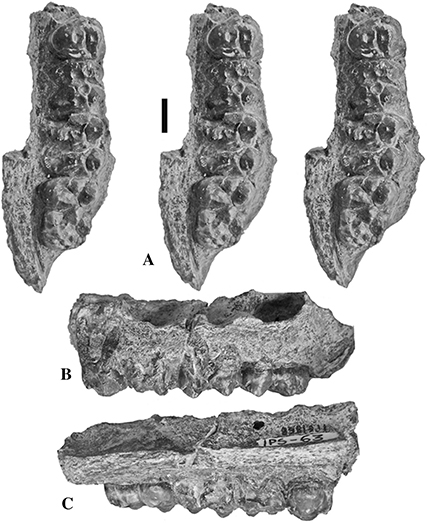 Fig. 14.— IPS 1868, left maxilla containing P4/-M3/, holotype of Retroporcus matritensis nov. comb. (Golpe-Posse, 1972) from Puente de Vallecas, Spain, A) stereo triplet of the occlusal view, B) buccal view, C) lingual view (scale: 10 mm). Fig. 14.— IPS 1868, left maxilla containing P4/-M3/, holotype of Retroporcus matritensis nov. comb. (Golpe-Posse, 1972) from Puente de Vallecas, Spain, A) stereo triplet of the occlusal view, B) buccal view, C) lingual view (scale: 10 mm).
|
|
The isolated right M2/ (MHNT PAL 2012.0.198) is deeply worn. It shows four main cusps with a strong median accessory cusplet,
a weaker anterior cusplet and a weak posterior cusplet. It has strong lingual flare, but the buccal surface of the tooth is
almost vertical.
|
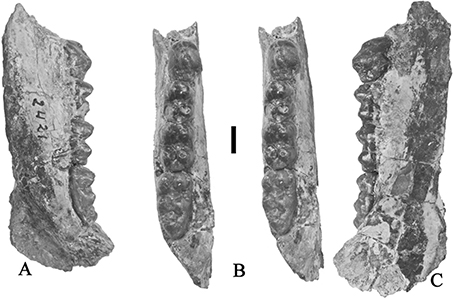 Fig. 15.— Retroporcus matritensis nov. comb. (Golpe-Posse, 1972) from Puente de Vallecas, the type locality. MSIM 2421, right mandible with p/4-m/3, A) lingual, B) stereo occlusal, C) buccal
views (scale: 10 mm). Fig. 15.— Retroporcus matritensis nov. comb. (Golpe-Posse, 1972) from Puente de Vallecas, the type locality. MSIM 2421, right mandible with p/4-m/3, A) lingual, B) stereo occlusal, C) buccal
views (scale: 10 mm).
|
|
MHNT PAL 2012.0.194 is a damaged right maxilla set in a plinth of plaster-of-paris. It contains the P4/ and M2/ in the bone,
but the M3/ is isolated behind. The roots of the P3/ and M1/ are partly preserved. The P3/ has three roots, the M1/ has four.
The P4/ is moderately worn, showing large dentine islands at the apices of both the buccal and the lingual cusps, surrounded
by thick enamel walls. The mesial and distal cingula are well developed. The M2/ is deeply worn and has lost part of the hypocone.
The mesial cingulum is broad, and there is a well developed basal pillar in the buccal end of the median transverse valley.
|
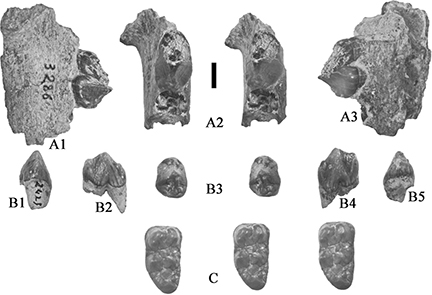 Fig. 16.— Lower teeth of Retroporcus matritensis nov. comb. (Golpe-Posse, 1972) from Puente de Vallecas, the type locality. A) MSIM 3286, left mandible fragment with p/3, A1) lingual, A2) stereo occlusal,
and A3) buccal views; B) MSIM 2421, right p/4, B1) mesial, B2) buccal, B3) stereo occlusal, B4) lingual, and B5) distal views;
C) MSIM 2423, right m/3, stereo occlusal views (scale: 10 mm). Fig. 16.— Lower teeth of Retroporcus matritensis nov. comb. (Golpe-Posse, 1972) from Puente de Vallecas, the type locality. A) MSIM 3286, left mandible fragment with p/3, A1) lingual, A2) stereo occlusal,
and A3) buccal views; B) MSIM 2421, right p/4, B1) mesial, B2) buccal, B3) stereo occlusal, B4) lingual, and B5) distal views;
C) MSIM 2423, right m/3, stereo occlusal views (scale: 10 mm).
|
|
The M3/ is moderately worn, with the talon in a position lingual to the midline of the crown. This makes the lingual surface
of the tooth almost straight from front to back, whereas the buccal surface curves markedly towards the rear. Most of the
paracone is broken off, and there is dentine exposed on the protocone. The talon complex is mesio-distally narrow.
In Paris there is a left mandible fragment containing a worn m/3 (MNHN VAS 16) which has written on it in the handwriting
of E. Lartet, the words Villefranche d’A. In the m/3 there is dentine exposed on all the main cusps and accessory cusplets,
as well as on the hypoconulid. The enamel surrounding the enamel islands is thick, giving rise to rounded cusps with small
indentations where the Fürchen are preserved (Hünermann, 1968). The distal root is inclined distally but is oriented almost vertically in the jaw.
|
 Fig. 17.— Upper teeth of Retroporcus matritensis nov. comb. (Golpe-Posse, 1972) from Puente de Vallecas, the type locality. A) MSIM 2642, left P4/, stereo occlusal view; B) MSIM 2680, left maxilla fragment
with P4/-M1/, stereo occlusal view; C) MSIM 2424, left M3/, stereo occlusal view; D) MSIM 2422, right M2/, stereo occlusal
view (scale: 10 mm). Fig. 17.— Upper teeth of Retroporcus matritensis nov. comb. (Golpe-Posse, 1972) from Puente de Vallecas, the type locality. A) MSIM 2642, left P4/, stereo occlusal view; B) MSIM 2680, left maxilla fragment
with P4/-M1/, stereo occlusal view; C) MSIM 2424, left M3/, stereo occlusal view; D) MSIM 2422, right M2/, stereo occlusal
view (scale: 10 mm).
|
|
Simorre (Rajegats):
MNHN Si 319 is a left maxilla fragment with the canine alveolus, the P1/, P2/ and P3/. There are short diastemata (5 mm) between
the canine alveolus and the P1/, and between the latter tooth and the P2/ (specimen Si 319). The canine alveolus reveals that
the tooth possessed a short root, and that it had a highly compressed section. The teeth in the maxilla are moderately (P1/-P2/)
to heavily worn (P3/). The P1/ has two stout roots, the anterior one smaller than the posterior one. The main cusp lies above
the gap between the two roots. The precrista descends from the apex of the main cusp towards the low mesial cusplet which
has broken off leaving low buccal and lingual cingular crests. The distal crista bifurcates distally, with one crest slanting
disto-buccal, the other distally. There is a low lingual cingulum walling off a shallow disto-lingual basin. The P2/ is a
larger version of the P1/ with the main structures enhanced. The disto-buccal crista swells into a low cusplet. The P3/ differs
from the preceding teeth by being considerably larger and taller, and in possessing three roots. The crown is heavily worn,
but it is possible to make out a well formed mesial cusplet at the end of the mesial crista, from which cingular crests run
rootwards buccally and lingually. A remnant of the lingual cingulum remains, but the linguo-distal cusplet has been worn away.
In lateral view, it is possible to observe a distinct vertical offset between the cervix of the P1/-P2/ on the one hand, and
the P3/ on the other, the distal part of the crown of P2/ touching the root of the P3/ about 3.5 mm beneath the mesial cusplet
of the P3/ and about 8 mm beneath its occlusal surface. This specimen is similar in all respects to Conohyus matritensis from Villefranche d’Astarac.
|
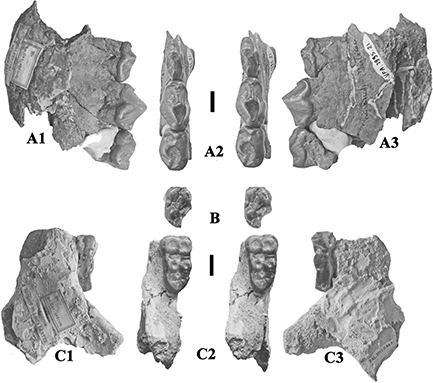 Fig. 18.— Retroporcus matritensis nov. comb. (Golpe-Posse, 1972) from Villefranche d’Astarac, France. A) MHNT PAL 2012.0.197, left mandible with p/2-p/3 and isolated p/4, A1) buccal, A2)
stereo occlusal, and A3) lingual views, B) MHNT PAL 2012.0.195, left m/1, stereo occlusal view, C) MHNT PAL 2011.0.84.3, left
mandible fragment containing m/3, C1) buccal, C2) stereo occlusal and C3) lingual views (scales: 10 mm). Fig. 18.— Retroporcus matritensis nov. comb. (Golpe-Posse, 1972) from Villefranche d’Astarac, France. A) MHNT PAL 2012.0.197, left mandible with p/2-p/3 and isolated p/4, A1) buccal, A2)
stereo occlusal, and A3) lingual views, B) MHNT PAL 2012.0.195, left m/1, stereo occlusal view, C) MHNT PAL 2011.0.84.3, left
mandible fragment containing m/3, C1) buccal, C2) stereo occlusal and C3) lingual views (scales: 10 mm).
|
|
MNHN Si 320 is interpreted to be an upper right P1/, but the crown is slightly damaged, such that it might represent a deciduous
rather than a permanent tooth.
MNHN Si 321 is a left P1/ similar to that in the maxilla described above.
|
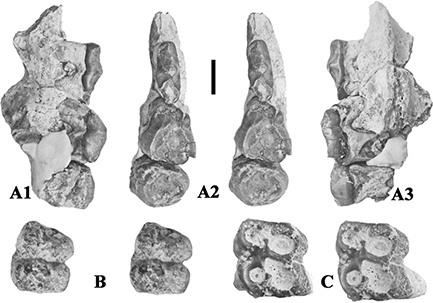 Fig. 19.— Retroporcus matritensis nov. comb. (Golpe-Posse, 1972) from Villefranche d’Astarac, France. A) MHNT PAL 2012.0.196, right maxilla containing P2/-P3/ and isolated P4/, A1) buccal,
A2) stereo occlusal and A3) lingual views, B) MHNT PAL 2012.0.199, right D4/, stereo occlusal view, C) MHNT PAL 2012.0.198,
right M2/, stereo occlusal view (scale: 10 mm). Fig. 19.— Retroporcus matritensis nov. comb. (Golpe-Posse, 1972) from Villefranche d’Astarac, France. A) MHNT PAL 2012.0.196, right maxilla containing P2/-P3/ and isolated P4/, A1) buccal,
A2) stereo occlusal and A3) lingual views, B) MHNT PAL 2012.0.199, right D4/, stereo occlusal view, C) MHNT PAL 2012.0.198,
right M2/, stereo occlusal view (scale: 10 mm).
|
|
A right P3/ in the sample (SML 463) is in light wear.
The unworn P4/, MNHN SML 78, has a buccal main cusp which is subdivided at its apex into two points, a large mesial one, and
a smaller, lower one immediately behind it. The lingual main cusp (protocone) is lower than the buccal one, and sends a precrista
towards the midline of the mesial margin of the tooth but it does not join the mesial cingulum. The paracone sends two crests
anteriorly, one directly towards a low cusplet at the buccal end of the mesial cingulum (parastyle), the other from its lingual
surface anteriorly towards the midline of the tooth.The posterior cingulum is beaded and swells in the end of the sagittal
valley to form a low distal accessory cusplet. The cingulum weakens buccally but is continuous from front to rear. There is
no sign of the presence of cusplets in the sagittal valley. Both the buccal and lingual surfaces of the crown are strongly
flared, making the tooth considerably broader than long.
|
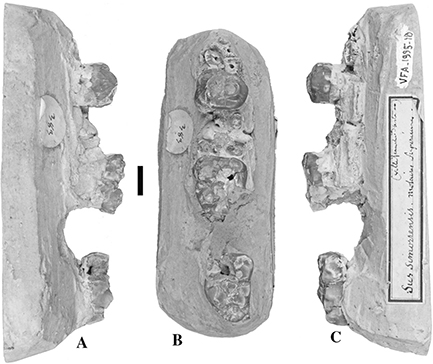 Fig. 20.— Retroporcus matritensis nov. comb. (Golpe-Posse, 1972) from Villefranche d’Astarac, France. MHNT PAL 2012.0.194, plaster plinth holding a right maxilla containing roots of P3/,
crowns of P4/ and M2/, plus an isolated M3/, A) buccal, B) occlusal, and C) lingual views (scale: 10 mm). Note that the M3/
should be up against the M2/ and not separated from it. Fig. 20.— Retroporcus matritensis nov. comb. (Golpe-Posse, 1972) from Villefranche d’Astarac, France. MHNT PAL 2012.0.194, plaster plinth holding a right maxilla containing roots of P3/,
crowns of P4/ and M2/, plus an isolated M3/, A) buccal, B) occlusal, and C) lingual views (scale: 10 mm). Note that the M3/
should be up against the M2/ and not separated from it.
|
|
The P4/s in the sample (MNHN SML 234, 337) are quite variable in dimensions, but all show the same basic morphology as the
germ described above.
MNHN SML 324 is a worn right D4/. It is constructed along the lines of a permanent molar, but has a more trapezoidal occlusal
outline, and thinner enamel.
|
 Fig. 21.— Retroporcus matritensis nov. comb. (Golpe-Posse, 1972) from Villefranche d’Astarac, France housed in the Muséum National d’Histoire Naturelle, Paris. A-C) MNHN VAS 16, left mandible
fragment containing m/3, A) buccal, B) stereo occlusal, C) lingual, and D) and posterior views (scale: 10 mm). Note the vertical
distal root beneath the talonid of the m/3. Fig. 21.— Retroporcus matritensis nov. comb. (Golpe-Posse, 1972) from Villefranche d’Astarac, France housed in the Muséum National d’Histoire Naturelle, Paris. A-C) MNHN VAS 16, left mandible
fragment containing m/3, A) buccal, B) stereo occlusal, C) lingual, and D) and posterior views (scale: 10 mm). Note the vertical
distal root beneath the talonid of the m/3.
|
|
An upper third molar from Simorre (MNHN SM 290) is broad distally due to relatively great development of the structures lingual
to the hypocone, and the poor development of the talon complex.
|
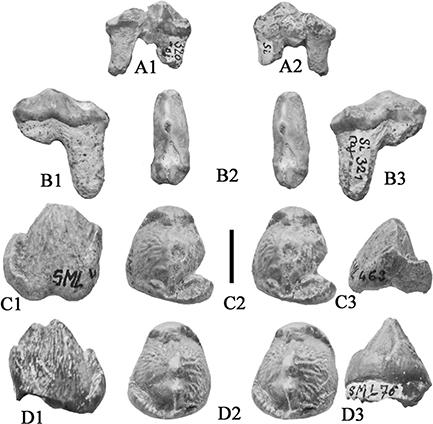 Fig. 22.— Tetraconodont anterior upper premolars from Simorre, France, housed in the Muséum National d’Histoire Naturelle, Paris.
A) SML 320, right P1/, Retroporcus matritensis nov. comb. A1) lingual, A2) buccal, B) SML 321, left P1/, Retroporcus matritensis, B1) lingual, B2) stereo occlusal, B3) buccal, C) SML 463, right P3/, Retroporcus matritensis C1) buccal, C2) stereo occlusal, C3) distal, D) SML 76, left P3/, Conohyus simorrensis, D1) buccal, D2) stereo occlusal, D3) distal (scale: 10 mm). Fig. 22.— Tetraconodont anterior upper premolars from Simorre, France, housed in the Muséum National d’Histoire Naturelle, Paris.
A) SML 320, right P1/, Retroporcus matritensis nov. comb. A1) lingual, A2) buccal, B) SML 321, left P1/, Retroporcus matritensis, B1) lingual, B2) stereo occlusal, B3) buccal, C) SML 463, right P3/, Retroporcus matritensis C1) buccal, C2) stereo occlusal, C3) distal, D) SML 76, left P3/, Conohyus simorrensis, D1) buccal, D2) stereo occlusal, D3) distal (scale: 10 mm).
|
|
Diversity of tetraconodonts at Villefranche d’AstaracTOP
The first conclusion is that tetraconodont fossils from Villefranche d’Astarac represent two distinct taxa: 1) Conohyus simorrensis, in which the p/2 is somewhat sectorial, is positioned at the same level as the p/3, and with the occlusal surface of the
talonid of the m/3 twisted with respect to that of the two anterior lophids, the talonid is slightly buccally positioned with
respect to the midline of the tooth, and the distal root of m/3 is slightly inclined buccally; 2) a second taxon in which
the p/2 is more bunodont and is offset vertically with respect to the p/3, the occlusal surface of the talonid of m/3 is not
twisted, the talonid of m/3 is more or less in line with the midline of the tooth, and the distal root of the m/3 is oriented
almost vertically in the jaw. Both these taxa differ markedly from the Steinheim tetraconodont (Chaeropotamus steinheimensis of Fraas, 1870) which has very buccally inclined distal roots in the m/3, and in which the talonid of the m/3 is in line with the buccal
cusps of the two anterior lophids, the p/2 is bunodont with swollen and ridged disto-buccal complexes, and the p/2 is not
vertically offset with respect to p/3.
The second conclusion is that the second tetracondont species from Villefranche d’Astarac is morphometrically similar to Retorporcus matritensis nov. comb. from Puente de Vallecas, Spain and Göriach, Austria (Conohyus simorrensis goeriachensis of Van der Made, 1989). A morphologically similar but smaller taxon, Retroporcus complutensis nov. gen. nov. sp. (the type species of the genus) occurs at Somosaguas (MN 5, Spain) (Van der Made & Salesa, 2004).
|
 Fig. 23.— Tetraconodont upper fourth premolars from Simorre, France, housed in the Muséum National d’Histoire Naturelle, Paris. A)
SML 464, right P4/, Conohyus simorrensis, A1) stereo occlusal, A2) mesial, B) SML 78, left P4/, Retroporcus matritensis nov. comb. stereo occlusal view, C) SML 337, right P4/, Retroporcus matritensis, C1) stereo occlusal, C2) mesial, D) SML 234, left P4/, Retroporcus matritensis, stereo occlusal view, E) SML 37, right P4, E1) stereo occlusal, E2, distal (scale: 10 mm). Fig. 23.— Tetraconodont upper fourth premolars from Simorre, France, housed in the Muséum National d’Histoire Naturelle, Paris. A)
SML 464, right P4/, Conohyus simorrensis, A1) stereo occlusal, A2) mesial, B) SML 78, left P4/, Retroporcus matritensis nov. comb. stereo occlusal view, C) SML 337, right P4/, Retroporcus matritensis, C1) stereo occlusal, C2) mesial, D) SML 234, left P4/, Retroporcus matritensis, stereo occlusal view, E) SML 37, right P4, E1) stereo occlusal, E2, distal (scale: 10 mm).
|
|
Discussion and ConclusionsTOP
The discovery of the long “missing” fossil collections from Simorre and Villefranche d’Astarac studied by Lartet (1851) is an important event in suid palaeontology. Details of the events culminating in the “loss” of the collections are scanty,
but it is sure that by the end of the 19th Century they had been misplaced and were considered lost by Stehlin (1899–1900) who examined collections in many museums
throughout Western Europe and described specimens that had not been seen for many years, including fossils from Bonnefond,
Le Fousseret and St Gaudens (Valentine). But the important material from Villefranche d’Astarac, which was the main basis
for Lartet’s (1851) decision to erect the species Sus simorrensis, evaded him. The only evidence of this material he reported seeing (Stehlin, 1899–1900, p. 45) was a cast of MHNT PAL 2012.0.206 kept in the university museum in Bologna, Italy, which he later wrote represented
a fossil from Le Fousseret (Stehlin, 1899–1900, p. 140).
|
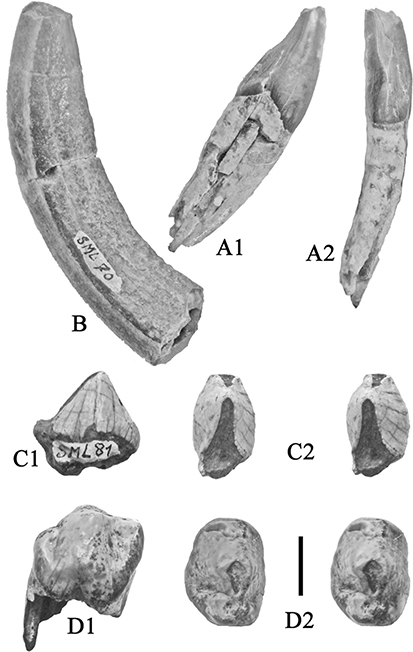 Fig. 24.— Tetraconodont lower incisor, lower canine and lower premolars from Simorre, France, housed in the Muséum National d’Histoire
Naturelle, Paris. A) SML 683, right i/2, Conohyus sp., A1) distal, A2, lingual, B) SML 70, left lower canine, Retroporcus matritensis nov. comb. distal view, C) SML 81, left d/3, Conohyus simorrensis, C1) lingual, C2) stereo oclusal, D) SML 39, left p/4, Conohyus simorrensis, D1) buccal, D2) stereo occlusal (scale: 10 mm). Note the absence of a ridge of cementum on the distal, enamel-free surface
of the canine. Fig. 24.— Tetraconodont lower incisor, lower canine and lower premolars from Simorre, France, housed in the Muséum National d’Histoire
Naturelle, Paris. A) SML 683, right i/2, Conohyus sp., A1) distal, A2, lingual, B) SML 70, left lower canine, Retroporcus matritensis nov. comb. distal view, C) SML 81, left d/3, Conohyus simorrensis, C1) lingual, C2) stereo oclusal, D) SML 39, left p/4, Conohyus simorrensis, D1) buccal, D2) stereo occlusal (scale: 10 mm). Note the absence of a ridge of cementum on the distal, enamel-free surface
of the canine.
|
|
Selecting a lectotype from the sample of bunodont suids from Villefranche d’Astarac is not a simple affair. Our examination
of the fossils reveals that there are two morphotypes of bunodont suids at Villefranche d’Astarac, both with enlarged posterior
premolars and reduced anterior ones. One is comparable in morphology to the material traditionally thought of as representing
Conohyus simorrensis, based on the left and right mandibles from Le Fousseret (MHNT PAL 2012.0.206 and MNHN HGP 16). The other consists of specimens
in which the anterior premolars are more bunodont than the somewhat sectorial ones that occur in Conohyus simorrensis, and which are vertically offset in the jaw relative to the P3/ and p/3. The latter morphotype occurs at Puente de Vallecas,
Spain (named Hyotherium soemmeringi matritensis by Golpe Posse, 1972) and Göriach, Austria (Hofmann, 1893), the latter collection named Conohyus simorrensis goeriachensis by Van der Made, 1989. A smaller species related to the latter morphotype occurs at Somosaguas, Spain (Van der Made & Salesa, 2004) among other sites and is here named Retroporcus complutensis nov. gen. nov. sp.
|
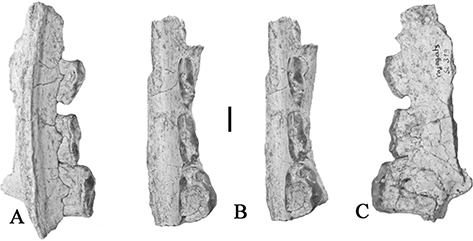 Fig. 25.— MNHN Si 319, left maxilla of Retroporcus matritensis nov. comb. containing P1/-P3/, from Simorre, France, housed in the Muséum National d’Histoire Naturelle, Paris, A) lingual,
B) stereo occlusal, C) buccal views (scale: 10 mm). Fig. 25.— MNHN Si 319, left maxilla of Retroporcus matritensis nov. comb. containing P1/-P3/, from Simorre, France, housed in the Muséum National d’Histoire Naturelle, Paris, A) lingual,
B) stereo occlusal, C) buccal views (scale: 10 mm).
|
|
Several authors (Hünermann, 1968; Chen, 1984; Thenius, 1956) were uneasy with the fact that the type material of Sus simorrensis from France had never been described in detail or measured. Various proposals were put forward to nominate alternative specimens
from either Simorre or Le Fousseret as lectotypes or neotypes, but these were never formalised. As it happens, even though
the specimens alluded to by Van der Made (1989, 1998) include partial tooth rows, they did not preserve the most diagnostic teeth which are the anterior premolars and the third
molars complete with their roots.
To complicate matters, fossils labelled Le Fousseret, including a palate erroneously said to be from the site, were used by
Van der Made (1989) as a basis for separating the Göriach sample from the French one, in the process of which he named Conohyus simorrensis goeriachensis. The palate (MNHN HGP 17) is here considered to be the long lost snout from Valentine, the type specimen of Sus valentini described by Filhol (1882). Thus the concept of the species Conohyus simorrensis that Van der Made (1989) had in mind, was based partly on fossils that fall outside the range of morphometric variation of the assemblage from Villefranche
d’Astarac used by Lartet (1851) when naming the species.
|
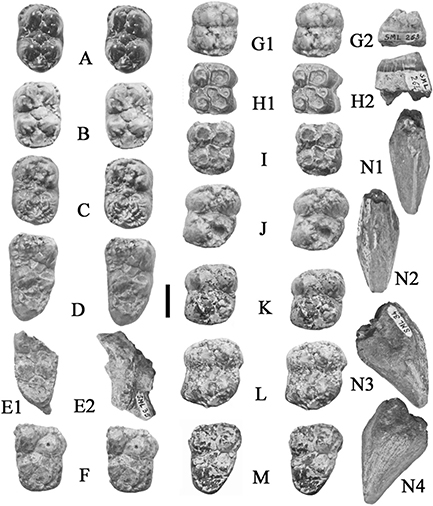 Fig. 26.— Tetraconodont upper canine and upper and lower molars from Simorre, France, housed in the Muséum National d’Histoire Naturelle,
Paris. A) SML 1144, left m/2, Conohyus sp., stereo occlusal, B) SML 77, right m/2, Conohyus sp., stereo occlusal, C) SML 73, left m/2, Conohyus sp., stereo occlusal, D) SML 74, left m/3, Conohyus simorrensis, stereo occlusal, E) SML 35, right m/3 fragment, Conohyus simorrensis, E1) occlusal, E2, buccal view, F) SML 290, right M3/, Retroporcus matritensis nov. comb. stereo occlusal, G) 263, left M1/, Conohyus sp., stereo occlusal, H) SML 264, right M1/, Conohyus sp., stereo occlusal, I) Si 324, right M1/, Retroporcus matritensis, stereo occlusal J) SML 79, left M2/, Conohyus sp., stereo occlusal, K) SML 57, left M2/, Conohyus simorrensis, stereo occlusal, L) SML 75, left M2/, Conohyus sp., stereo occlusal, M) SML 36, left M3/, Conohyus simorrensis, stereo occlusal, N) SML 34, right upper canine, Conohyus sp., N1) posterior, N2) anterior, N3) buccal, N4) lingual views (scale: 10 mm). Fig. 26.— Tetraconodont upper canine and upper and lower molars from Simorre, France, housed in the Muséum National d’Histoire Naturelle,
Paris. A) SML 1144, left m/2, Conohyus sp., stereo occlusal, B) SML 77, right m/2, Conohyus sp., stereo occlusal, C) SML 73, left m/2, Conohyus sp., stereo occlusal, D) SML 74, left m/3, Conohyus simorrensis, stereo occlusal, E) SML 35, right m/3 fragment, Conohyus simorrensis, E1) occlusal, E2, buccal view, F) SML 290, right M3/, Retroporcus matritensis nov. comb. stereo occlusal, G) 263, left M1/, Conohyus sp., stereo occlusal, H) SML 264, right M1/, Conohyus sp., stereo occlusal, I) Si 324, right M1/, Retroporcus matritensis, stereo occlusal J) SML 79, left M2/, Conohyus sp., stereo occlusal, K) SML 57, left M2/, Conohyus simorrensis, stereo occlusal, L) SML 75, left M2/, Conohyus sp., stereo occlusal, M) SML 36, left M3/, Conohyus simorrensis, stereo occlusal, N) SML 34, right upper canine, Conohyus sp., N1) posterior, N2) anterior, N3) buccal, N4) lingual views (scale: 10 mm).
|
|
In order to respect the concept of Conohyus simorrensis articulated by Stehlin (1899–1900) who effectively provided the first detailed descriptions and illustrations of the species,
we have selected as lectotype, a lower jaw from Villefranche d’Astarac, comprised of three fragments, two of which have been
stuck together with plaster-of-paris. This specimen agrees in all essential morphological details with, but is slightly smaller
than the mandible from Le Fousseret (NHMT PAL 2011.0.84.2) upon which Stehlin (1899–1900) based his concept of the species,
and which has been provisionally proposed on various occasions as a lectotype, but never formally designated as such (Chen,
1984; Van der Made & Morales, 2003). The second tetraconodont species from Villefranche d’Astarac already has a name in Hyotherium simorrense matritensis (Golpe-Posse, 1972) of which the type specimen is from Puente de Vallecas, Spain. We here revise the rank of the taxon and classify it in a
new genus (Retroporcus, type species Retroporcus complutensis) as the combination Retroporcus matritensis nov. comb. (Golpe-Posse, 1972).
|
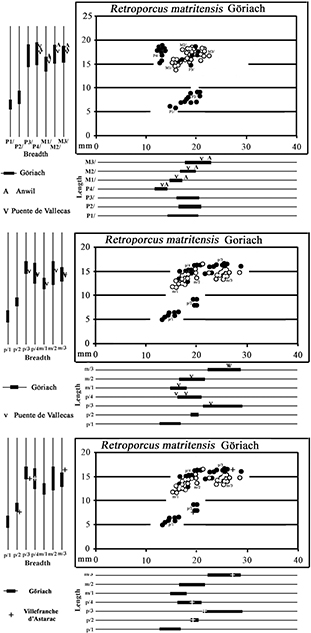 Fig. 27.— Bivariate and univariate plots of the upper (top) and lower cheek teeth (middle and bottom) of Retroporcus matritensis nov. comb. from Puente de Vallecas, Anwil (Engesser, 1972) and Göriach. The circles are specimens from Göriach, Austria, which is a reference assemblage for comparison of fossils
from other localities defined by letters of the alphabet in the univariate plots Puente de Vallecas (V symbol), Villafranche
d’Astarac (+ symbol) (premolars – solid symbols; molars – hollow symbols). Fig. 27.— Bivariate and univariate plots of the upper (top) and lower cheek teeth (middle and bottom) of Retroporcus matritensis nov. comb. from Puente de Vallecas, Anwil (Engesser, 1972) and Göriach. The circles are specimens from Göriach, Austria, which is a reference assemblage for comparison of fossils
from other localities defined by letters of the alphabet in the univariate plots Puente de Vallecas (V symbol), Villafranche
d’Astarac (+ symbol) (premolars – solid symbols; molars – hollow symbols).
|
|
Finally, we stress the fact that the rediscovery of the Lartet (1851) collection of suids from Villefranche d’Astarac and Simorre will necessitate a revision of the European and Asian tetraconodonts,
as it is now clear that the fossils previously attributed to the genus Conohyus comprise at least two genera. This paper revises two of the genera represented in Europe (Conohyus and Retroporcus nov. gen.). The other genera (Parachleuastochoerus, Sivachoerus) will be revised in a forthcoming article.
ACKNOWLEDGEMENTSTOP
Access to fossil collections was accorded by many colleagues: Herbert Lutz and Thomas Engel at the Naturhistorisches Museum,
Mainz, Oliver Sandrock and Eric Milson at the Hessisches Landesmuseum, Darmstadt. We have been helped by curators in London
(Andy Currant, Pip Brewer, Emma Bernard), München (Gertrud Rössner), Stuttgart (Reinhard Ziegler), Vienna (Ursula Göhlich),
Graz (Martin Gross), Madrid (Jorge Morales, Patricia Perez), Lyon (Abel Prieur, Emmanuel Robert, Didier Berthet), Toulouse
(Francis Duranton), Basel (Loïc Costeur, Martin Schneider), Paris (Christine Argot, Claire Sagne), Universidad Complutense
(Manuel Hernandez), Museo San Isidro, Madrid (Mercedes Gamazo), Instituto Geologico y Minero de España (Alfonso Arribas, Guiomar
Garrido), Institut Català de Paleontologia Miquel Crusafont, Sabadell (Salvador Moyà-Solà, Dani DeMiguel, Laura Celia) – we
thank them all. Carlo Sarti (University of Bologna) kindly sent images of the cast of the mandible from Le Fousseret.
ReferencesTOP
| ○ |
Antunes, M.T., Ginsburg, L. & Mein, P. (1983). Mammifères miocènes de Azambujeira, niveau inférieur (Santarém, Portugal).
Ciencias da Terra (UNL), Lisboa, 7: 161–186.
|
| ○ |
Azanza, B. (1986). Estudio geologico y paleontologico del Mioceno del sector oeste de la Comarca de Borja. Cuadernos de Estudios
Borjanos, 7–8: 63–126.
|
| ○ |
Blainville, H.M.D. de (1847). Ostéographie ou description iconographique comparé du squelette et du système dentaire des cinq
classes d’animaux vertébrés récents et fossiles, pour servir de base à la Zoologie et à la Geologie. 4, AA, Sur les Hippopotames et les Cochons, Paris, Bertrand, 248 pp. |
| ○ |
Chen, Guanfang (1984). Suidae and Tayassuidae (Artiodactyla, Mammalia) from the Miocene of Steinheim a. A. (Germany). Palaeontographica,
184(1–4): 79–83.
|
| ○ |
Engesser, B. (1972). Die obermiocäne Säugetierfauna von Anwil (Baselland). Tätigkeitsberichte der naturforschenden Gesellschaft
Baselland, 28: 37–363.
|
| ○ |
Falconer, H. & Cautley, P.T. (1847). Fauna antiqua sivalensis. London, 107 pp. |
| ○ |
Filhol, H. (1882). Note relative à une nouvelle espèce de Sus fossile trouvée dans les argiles à Dinotherium de Valentine (Haute-Garonne). Bulletin de la Société philomathique de Paris, Séries 7, 6: 123–124.
|
| ○ |
Fortelius, M., Van der Made, J. & Bernor, R. (1996). Middle and Late Miocene Suoidea of Central Europe and the Eastern Mediterranean:
Evolution, Biogeography and Paleoecology. In: R.L. Bernor, V. Fahlbusch & H.W. Mittmann (eds.) The Evolution of Western Eurasian Neogene Mammal Faunas. Columbia University Press, New York, 348–377.
|
| ○ |
Fraas, O. (1870). Beiträge zur Fauna von Steinheim. Mit Rüchsicht auf die miocenen Säugetier- und vogelreste des Steinheimer
Beckens. Jahreshefte der Vereinigung Vaterländische Naturkunde Würtembergs, 26: 145–306.
|
| ○ |
Ginsburg, L. (1977). Sur la répartition stratigraphique de Conohyus simorrensis (Suidae, Artiodactyla, Mammalia) dans le Miocène européen. Comptes rendus sommaries de la Société géologique de France, 1977(4):
203–205.
|
| ○ |
Golpe-Posse, J.M. (1972). Suiformes del Tertiario Español y sus yacimientos. Paleontologia y Evolucion, 2: 1–198. |
| ○ |
Gray, J.E. (1821). On the natural arrangement of vertebrose animals. London Medical Repository, 15(1): 296–310. |
| ○ |
Hofmann, A. (1893). Die Fauna von Göriach. Abhandlungen der k.k. geologische Reischsanstalt, 15: 1–87. |
| ○ |
Hünermann, K. A. (1968). Die Suidae (Mammalia, Artiodactyla) aus den Dinotheriensanden (Unterpliozän = Pont) Rheinhessens
(Südwestdeutschland). Schweizerische Paläontologische Abhandlungen, 86: 1–96.
|
| ○ |
Kaup, J.J. (1833). Description d’ossements fossiles de Mammifères, 2. Darmstadt, J.G. Heyer, 31 pp. |
| ○ |
Kaup, J.J. (1859). Beiträge zur näheren Kenntnis der urweltlichen Säugethiere, Volume 4, Darmstadt, 31 pp. |
| ○ |
Lartet, E. (1851). Notice sur la Colline de Sansan, suivi d’une récapitulation des diverses espèces d’animaux vertébrés fossiles,
trouvés soit à Sansan, soit dans d’autres gisements du terrain tertiaire miocène dans le bassin sous-pyrénéen. J-A Portes,
Auch, 65 pp.
|
| ○ |
Lydekker, R. (1876). Fossil mammalian faunae of India and Burma. Records of the Geological Survey of India, 9: 86–106. |
| ○ |
Lydekker, R. (1884). Indian Tertiary and Post-Tertiary Vertebrata: Siwalik and Narbada Bunodont Suina. Memoirs of the Geological
Survey of India, Palaeontologica indica, (10) 3 (2): 35–104.
|
| ○ |
Mein, P. (1986). Chronological succession of hominoids in the European Neogene In: Else J.G. & Lee P.C. (eds.) Primate Evolution,
Cambridge, Cambridge University Press, Vol 1, pp. 59–82.
|
| ○ |
Morales, J., & Soria, D. (1985). Carnivoros y Artiodactilos de la Provincia de Madrid. In: M.T. Alberdi (Co-ord.) Geologia
y Paleontologia del Tertiario continental de la Provincia de Madrid. Instituto de Geologia (CSIC), Madrid, 81–92. |
| ○ |
Petronijevic, Z.M. (1967). Srednjomiocenska I donjosarmatska (Stajerska) Fauna Sisara Srbije. Palaeontologia Jugoslavica,
7: 1–160.
|
| ○ |
Pickford, M. (1988). Revision of the Miocene Suidae of the Indian Subcontinent. Münchner Geowissenschaftliche Abhandlungen,
12: 1–91.
|
| ○ |
Pickford, M. (2013). Conohyus simorrensis (Lartet, 1851) (Suidae, Mammalia) from the Middle Miocene of Carpetana (Madrid, Spain). Spanish Journal of Paleontology, 28(1): 91–102.
|
| ○ |
Pilgrim, G.E. (1925). Presidential address to the Geological Section of the 12th Indian Science Congress. Proceedings of the 12th Indian Scientific Congress, pp. 200–218.
|
| ○ |
Pilgrim, G.E. (1926). The Fossil Suidae of India. Memoir of the Geological Survey of India, Palaeontologica indica, 8(4):
1–65.
|
| ○ |
Richard, M. (1946). Contribution à l’étude du Bassin d’Aquitaine – Les gisements de mammifères Tertiaires. Mémoires de la
Société géologique de France (nouvelle série) 32: 1–380.
|
| ○ |
Roman, F. (1907). Le Néogène continental dans le Basse Vallée du Tage (Rive droite). Commission du Service Géologique de Portugal,
Lisbonne, 1907: 1–78.
|
| ○ |
Stehlin, H.G. (1899–1900). Über die Geschichte des Suiden Gebisses. Abhandlungen der Schweizerischen Paläontologischen Gesellschaft,
Zürich, 26/27: 1–527.
|
| ○ |
Thenius, E. (1956). Die Suiden und Tayassuiden der Steirischen Tertiärs. Beiträge zur Kenntniss des Säugetierreste des steirischen
Tertiärs VIII. Sitzungsberichten der Österreichische Akademie der Wissenschaften in Wien, Mathematische-Naturwissenschaftliche
Klasse, Abteilung 1, 165: 337–382.
|
| ○ |
Van der Made, J. (1989). A Conohyus lineage (Suidae, Artiodactyla) from the Miocene of Europe. Revista Española de Paleontologia, 4: 19–28.
|
| ○ |
Van der Made, J. (1990). Iberian Suoidea. Paleontologia I Evoluciò, 23: 83–97. |
| ○ |
Van der Made, J. (1998). Biometrical trends in the Tetraconodontinae, a subfamily of pigs. Transactions of the Royal Society
of Edinburgh: Earth Sciences, 89: 199–225. http://dx.doi.org/10.1017/S0263593300007136 |
| ○ |
Van der Made, J. & Morales, J. (2003). The pig Conohyus simorrensis from the Upper Aragonian of Alhambra, Madrid, and a review of the distribution of European Conohyus. Estudios geologicos, 59: 303–312.
|
| ○ |
Van der Made, J. & Salesa, M. (2004). Early remains of the pig Conohyus simorrensis from the Middle Aragonian of Somosaguas near Madrid – its dispersal into Europe and evolution. Neues Jahrbuch für Geologie
und Paläontologie Abhandlungen. 233: 153–168.
|
| ○ |
Vitry, U. (1857). Académie Impériale des Sciences, Inscriptions et Belles-Lettres. Séance du 7 mai. Journal de Toulouse, 157:
1.
|