Early Pliocene fishes (Chondrichthyes, Osteichthyes) from Gran Canaria and Fuerteventura (Canary Islands, Spain)
Los peces (Chondrichthyes, Osteichthyes) del Plioceno inferior de Gran Canaria y Fuerteventura (Islas Canarias, España)
J.F. Betancort1, A. Lomoschitz2, J. Meco1
1 Departamento de Biología, Campus Universitario de Tafira, Universidad de Las Palmas de Gran Canaria, 35017 Las Palmas de Gran Canaria, Spain. Email: jbetancor@becarios.ulpgc.es; joaquinfrancisco.meco@ulpgc.es
2 Instituto de Oceanografía y Cambio Global (IOCAG), Campus Universitario de Tafira, Universidad de Las Palmas de Gran Canaria, 35017 Las Palmas de Gran Canaria, Spain. Email: alejandro.lomoschitz@ulpgc.es
| |
ABSTRACT
Fossil fish teeth are contained in marine deposits dated at ca 4.8 Ma found on the islands of Gran Canaria and Fuerteventura (Canary Islands, Spain). These islands, situated in the North
Atlantic Subtropical Gyre, can be considered a mid-way stopover point between the Caribbean Sea, with the Central American
Seaway about to close in this epoch, and the Mediterranean, in the first stage of its post-Messinian Gibraltar Seaway period.
Accordingly, there existed extensive pantropical communication, particularly for nektonic animals capable of travelling large
distances. In this paper, we present a number of fossil fishes, most of which are identified for the first time on the basis
of their teeth: the Chondrichthyes species Carcharocles megalodon, Parotodus benedeni, Cosmopolitodus hastalis, Isurus oxyrinchus, Carcharias cf. acutissima, Carcharhinus cf. leucas, Carcharhinus cf. priscus, Galeocerdo cf. aduncus, and the Osteichthyes species Archosargus cinctus, Labrodon pavimentatum, and Diodon scillae. Coincidences are observed between these ichthyofauna and specimens found in the Azores Islands, the Pacific coast of America
and the Mediterranean Sea.
Keywords: Biogeography;
Palaeoclimatology;
Pillow-lavas dating;
Subtropical North Atlantic;
Central American Seaway;
Mediterranean Flood.
|
| |
RESUMEN
Los dientes fósiles de peces están contenidos en depósitos marinos datados en ca 4.8 Ma de las Islas de Gran Canaria y Fuerteventura (Islas Canarias, España). Estas islas, situadas en el Giro subtropical
del Atlántico Norte, pueden considerarse una estación intermedia entre un Mar Caribe que estaba cercano al cierre del Paso
de América Central, y el Mediterráneo en su primera etapa tras la apertura post-Mesiniense de Gibraltar. Ello permitía una
comunicación pantropical para los animales del necton que eran capaces de desplazarse a grandes distancias. Se identifican
por primera vez los peces Chondrichthyes Carcharocles megalodon, Parotodus benedeni, Cosmopolitodus hastalis, Isurus oxyrinchus, Carcharias cf. acutissima, Carcharhinus cf. leucas, Carcharhinus cf. priscus, Galeocerdo cf. aduncus y las species de Osteichthyes Archosargus cinctus, Labrodon pavimentatum y Diodon scillae. Se ha observado coincidencia entre los ejemplares canarios y los encontrados en las Islas Azores, las costas pacíficas de
América y el mar Mediterráneo.
Palabras clave: Biogeografía;
Paleoclimatología;
Datación de Pillow-lavas;
Atlántico Norte Subtropical;
Paso de América Central;
Inundación del Mediterráneo.
|
IntroductionTOP
There is little documentation of Canary fossil fishes, with the exception of two publications from the 19th century: (1) the
study carried out by Cocchi (1864) in which he created the genus Pharyngodopilus and its two species Ph. canariensis and Ph. Africanus, catalogued in 1857 by the British Museum of Natural History and which formed part of the collection of Charles Lyell; and
(2) the study conducted by Rothpletz & Simonelli (1890), which includes Oxyrhina plicatilis, Ag., Oxyrhina sp., Galeocerdo cf. egertoni, Ag., Chrysophrys sp., Nummopalatus africanus, Cocchi and Diodon sigma Martin (Fig. 1). It should be noted that the age of the deposits which contained these fauna were ascribed to the Upper Miocene (Cocchi, 1864; Lyell, 1865) and were included in the Helvetian or Tortonian (Rothpletz & Simonelli, 1890).
|
 Fig. 1.—Diodon sigma, is one of the few remaining first fossil fishes from Las Palmas de Gran Canaria that were studied in 19th century. It remains in the Paläontologisches Museum München having survived two world wars. It was found at La Vista (Barranco Seco) by Rothpletz. This German geologist was visiting Las Palmas de Gran Canaria for health reasons according to an unpublished letter of his addressed to Dr. Chil who at the time (1879) was founding El Museo Canario. During his time on the island Rothpletz made an extensive collection of fossils (most of which have sadly been lost) which was studied by Simonelli and published by Rothpletz & Simonelli (1890). The deposits attributed to the Miocene have been recently dated at ca 4.80 Ma (Meco et al., 2015). Scale bar: 10 mm. Fig. 1.—Diodon sigma, is one of the few remaining first fossil fishes from Las Palmas de Gran Canaria that were studied in 19th century. It remains in the Paläontologisches Museum München having survived two world wars. It was found at La Vista (Barranco Seco) by Rothpletz. This German geologist was visiting Las Palmas de Gran Canaria for health reasons according to an unpublished letter of his addressed to Dr. Chil who at the time (1879) was founding El Museo Canario. During his time on the island Rothpletz made an extensive collection of fossils (most of which have sadly been lost) which was studied by Simonelli and published by Rothpletz & Simonelli (1890). The deposits attributed to the Miocene have been recently dated at ca 4.80 Ma (Meco et al., 2015). Scale bar: 10 mm.
|
|
The Canary Islands is a volcanic archipelago (Carracedo et al., 2002) and the marine deposits occur intercalated between lava-flows. Pillow-lavas are structures that are formed during the interaction
of lava and water. Accordingly, if a pillow-lava enters into contact with a marine deposit under formation, it indicates that
they both have the same age. This is the case of the Canary marine deposits containing the fossil fishes studied in this paper
that have a 40Ar/39Ar age of 4.80 ± 0.03 Ma in Gran Canaria (Meco et al., 2015) and a K/Ar age of 4.83 ± 0.10 Ma in Fuerteventura (Meco et al., 2007). This clearly places the marine deposits in the first half of the Early Pliocene which began 5.33 Ma ago and concluded around 3.60 Ma ago (Grandstein et al., 2004; Walker & Geissman, 2009).
On the basis of this information, we propose: (a) to present the list of fossil fishes contained in these dated deposits,
thereby contributing to the elaboration of a corrected biochronostratigraphy, and (b) to check if the fossil ichthyofauna
that have been found is consistent with the palaeoceanographic scenario in the area for the Early Pliocene. This scenario
is characterised principally by an open Central American Seaway (CAS) (Schmidt, 2007; Montes et al., 2012; Erkens, 2015; Meco et al., 2015; 2016), an open Gibraltar Seaway after the so-called Messinian crisis (Garcia-Castellanos et al., 2009) and warm pantropical waters (Meco et al., 2015) in the Pacific, Subtropical North Atlantic (Ávila et al., 2012) and Mediterranean (Pawellek et al., 2012).
Geological, palaeontological and stratigraphic settingTOP
The Canary Islands comprise seven main volcanic islands and several islets and are located in the Eastern Atlantic Ocean between
N 27° and N 30° latitudes, forming a chain that extends latitudinally about 450 km; with the easternmost point just over 100
km off the north-western African coast (Fig. 2). The islands have a complex geological history, with volcanic formations over 20 million years old (Carracedo et al., 2002), but they also include extensive sedimentary deposits (Meco & Stearns, 1981).
|
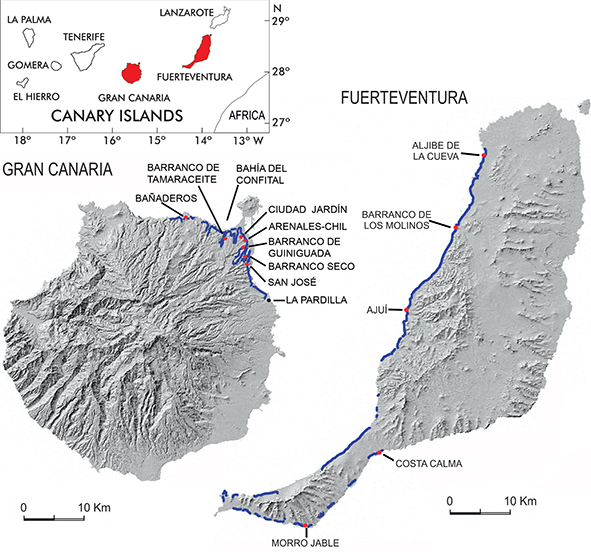 Fig. 2.—Early Pliocene sedimentary beds (lines in blue) with marine fossils of the Eastern Canary Islands (dated ca 4.80 Ma by Meco et al., 2015 and ca 4.83 Ma by Meco et al., 2007) and main sites (red dots) where the fossil marine fishes were found. Fig. 2.—Early Pliocene sedimentary beds (lines in blue) with marine fossils of the Eastern Canary Islands (dated ca 4.80 Ma by Meco et al., 2015 and ca 4.83 Ma by Meco et al., 2007) and main sites (red dots) where the fossil marine fishes were found.
|
|
They are composed of layers of two materials: (1) reddish conglomerate (rubefacted conglomerates as a result of Fe oxidation)
consisting of coarse gravel, cobble and medium to coarse size sand, and (2) grey to white sands, medium to fine in size, with
bioturbation structures, which constitute reference layers (or strata) in the local stratigraphy of each island. The original
conglomerates, comprised of cobbles and sands with a wide variety of fossils, were deposited on wave-cut platforms tens of
kilometres long. Subsequent withdrawal of the sea resulted in oxidation of pebbles and fossil remains because of contact with
freshwaters. These reddish sandy conglomerates as a whole are considered regional key layers, in both a stratigraphic and
palaeontological sense (Fritsch, 1867; Lomoschitz et al., 2011). All the fossil fish teeth that have been found in these deposits have a heavy layer of enamel with a dark orange colour.
The fauna which accompany the fossil fishes in Gran Canaria and Fuerteventura indicate ecological and climatic conditions
very different to those of today. The first information about invertebrate fossils of the Canary Islands was provided by Lyell (1865) and Rothpletz & Simonelli (1890). Detailed studies were later conducted of foraminifera (Anguita Virella & Ramírez del Pozo, 1974), of genus Strombus (=Perisitistrombus) (Meco, 1977), neogastropods (Meco, 1981), bivalves (Meco, 1982), bryozoans (Sendino & Taylor, 2014) and crustaceans (Betancort et al., 2014). The presence of fossil species belonging to the gastropod genus Persististrombus and Nerita, and the coral genus Siderastrea, and others, suggest climate conditions equivalent to those of present day tropical and sub-tropical regions, like the Caribbean
Sea (West Atlantic) or the Gulf of Guinea (East Atlantic) (Meco, 1977; Meco et al., 2007, 2015).
Stratigraphic sectionsTOP
The fossil fish specimens were found in sedimentary layers consisting of old coastal and marine deposits which outcrop along
over 130 km, if all the stretches are considered together (Fig. 2). As these layers show many lateral variations throughout the outcrops, three sections were chosen as representative of the
whole area: Barranco Seco and Tamaraceite sections from NE Gran Canaria Is. (Figs. 3 and 4) and Ajuí section from W Fuerteventura Is. (Figs. 3 and 5). The Barranco Seco locality provided the most numerous and the largest variety of fossil fishes, while the Tamaraceite and
Ajuí localities are characterised by the presence of lava flows which have allowed dating of the marine deposits at ca 4.8 Ma.
|
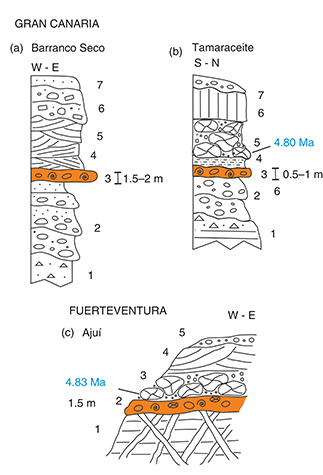 Fig. 3.—Schematic stratigraphic sections of sites on NE Gran Canaria and W Fuerteventura where the fossil marine fishes have been
found. A) Barranco Seco: 1 - Miocene phonolitic ignimbrite; 2 - Miocene alluvial fan; 3 - Early Pliocene marine deposit (reddish
conglomerate); 4 - Foreshore grey fine sand; 5 - Dune white sand; 6 - Pliocene alluvial fan; 7 - Colluvium. B) Tamaraceite
(ravine mouth): 1 - Miocene phonolitic lava flow and ignimbrite; 2 - Miocene alluvial fan; 3 - Early Pliocene marine deposit
(reddish conglomerate); 4 - White silty sand; 5 - Basaltic pillow lava 40Ar/39Ar dated ca 4.80 Ma (Meco et al., 2015); 6 - Basaltic lava flow; 7 - Colluvium. C) Ajuí: 1 - Pre-Miocene basaltic lava flows and dykes; 2 - Early Pliocene marine
deposit (reddish conglomerate); 3 - Basaltic pillow lava K/Ar dated ca 4.83 Ma (Meco et al., 2007); 4 - Pliocene dune; 5 - Colluvium. Fig. 3.—Schematic stratigraphic sections of sites on NE Gran Canaria and W Fuerteventura where the fossil marine fishes have been
found. A) Barranco Seco: 1 - Miocene phonolitic ignimbrite; 2 - Miocene alluvial fan; 3 - Early Pliocene marine deposit (reddish
conglomerate); 4 - Foreshore grey fine sand; 5 - Dune white sand; 6 - Pliocene alluvial fan; 7 - Colluvium. B) Tamaraceite
(ravine mouth): 1 - Miocene phonolitic lava flow and ignimbrite; 2 - Miocene alluvial fan; 3 - Early Pliocene marine deposit
(reddish conglomerate); 4 - White silty sand; 5 - Basaltic pillow lava 40Ar/39Ar dated ca 4.80 Ma (Meco et al., 2015); 6 - Basaltic lava flow; 7 - Colluvium. C) Ajuí: 1 - Pre-Miocene basaltic lava flows and dykes; 2 - Early Pliocene marine
deposit (reddish conglomerate); 3 - Basaltic pillow lava K/Ar dated ca 4.83 Ma (Meco et al., 2007); 4 - Pliocene dune; 5 - Colluvium.
|
|
|
 Fig. 4.—A. Outcrop at Barranco Seco site (NE Gran Canaria) where many fossil marine fishes have been collected: 1 - Phonolite non-welded
ignimbrite; 2 - Early Pliocene reddish conglomerate with fossils; 3 - foreshore grey fine sand; 4 - colluvium. B. Tooth of
Archosargus cinctus. Fig. 4.—A. Outcrop at Barranco Seco site (NE Gran Canaria) where many fossil marine fishes have been collected: 1 - Phonolite non-welded
ignimbrite; 2 - Early Pliocene reddish conglomerate with fossils; 3 - foreshore grey fine sand; 4 - colluvium. B. Tooth of
Archosargus cinctus.
|
|
|
 Fig. 5.—Outcrop at Ajuí site (W Fuerteventura): 1 - pre-Miocene volcanic and intrusive rocks; 2 - Early Pliocene marine deposit where some of the fossil specimens were found; 3a - basaltic pillow lava with a K/Ar age of ca 4.83 Ma (Meco et al., 2007); 3b - basaltic lava flow; 4 - Late Pliocene dune. Fig. 5.—Outcrop at Ajuí site (W Fuerteventura): 1 - pre-Miocene volcanic and intrusive rocks; 2 - Early Pliocene marine deposit where some of the fossil specimens were found; 3a - basaltic pillow lava with a K/Ar age of ca 4.83 Ma (Meco et al., 2007); 3b - basaltic lava flow; 4 - Late Pliocene dune.
|
|
Barranco Seco sectionTOP
Location: NE Gran Canaria Is., slope on Barranco Seco ravine.
Latitude: 28° 05′ 25.40″, longitude: 15° 25′ 40.25″, height: 85 m (apsl).
This section comprises seven units which are (bottom to top):
| 1. |
Phonolitic non-welded ignimbrite (>15 m).
Homogeneous in appearance, it is composed of thick layers of yellow to white rocks and corresponds to the Miocene of Gran
Canaria.
|
| 2. |
Heterogeneous sandy conglomerate of phonolitic pebbles (20–25 m).
This is a mixture of coarse gravel, pebbles and cobbles with a matrix of coarse sand and fine gravel. Their clasts are sub-angular
to sub-round in shape. It corresponds to a Miocene alluvial fan.
|
| 3. |
Reddish conglomerate with fossils (1.5–2 m).
The variable outcropping thickness of this layer is due to coastal erosion that occurred on an old phonolite cliff. A later
exposure to fresh water caused the red colour, which is common in weathering processes.
|
| 4. |
Grey sand with bioturbation structures (3–8 m). It is a deposit of medium to fine sand which originated in a foreshore environment. |
| 5. |
White sand dunes (5–10 m). Aeolianite composed of carbonate grains of fine to medium sand, with a characteristic cross-bedding
internal structure.
|
| 6. |
Heterogeneous sandy conglomerate of phonolitic and basaltic pebbles (20–25 m).
This is a mixture of coarse gravel with sand, cobbles and boulders, angular to sub-round in shape and slightly horizontally
layered. It is an alluvial fan of Pliocene age.
|
| 7. |
Colluvium. |
Tamaraceite sectionTOP
Location: NE Gran Canaria Is., slope on Tamaraceite ravine mouth.
Latitude: 28° 07″ 12.99″, longitude: 15° 27″ 30.64″, height: 97 m (apsl).
This section comprises seven units which are:
| 1. |
Phonolitic lava flows and non-welded ignimbrite (>15 m).
Homogeneous in appearance, it is composed of thick layers of yellow to white rocks and corresponds to the Miocene of Gran
Canaria.
|
| 2. |
Heterogeneous sandy conglomerate of phonolitic pebbles (20–25 m).
This is a mixture of coarse gravel, pebbles and cobbles with a matrix of coarse sand and fine gravel. Their clasts are sub-angular
to sub-round in shape. It corresponds to a Miocene alluvial fan.
|
| 3. |
Reddish conglomerate with fossils (0.5–1 m). |
| 4. |
White silty sand with fine parallel lamination (0.3–2 m). |
| 5. |
Basaltic pillow lavas and hyaloclastite (25 m) with a 40Ar/39Ar age of ca 4.8 Ma (Meco et al., 2015).
|
| 6. |
Basaltic lava flows (15 m). |
| 7. |
Colluvium. |
Ajuí sectionTOP
Location: W Fuerteventura Is., slope on Barranco de Ajuí ravine mouth.
Latitude: 28° 24″ 03.86″, longitude: 14° 09″ 21.20″, height: 7 m (apsl).
This section comprises five units which are:
| 1. |
Pre-Miocene volcanic, intrusive and sedimentary rocks. |
| 2. |
Reddish conglomerate with fossils (1.5 m). It is mainly composed of basaltic sub-rounded pebbles, gravels and cobbles with
a sandy matrix.
|
| 3. |
Basaltic pillow lavas and hyaloclastite (3 m). They come from olivine basaltic lava flows that erupted about 9 km inland.
This lava-flow has a K/Ar age of ca 4.83 Ma (Meco et al., 2007).
|
| 4. |
Pliocene white sand dunes (2 m). Aeolianite composed of carbonate grains of medium to fine sand, with a characteristic cross-bedding
internal structure.
|
| 5. |
Colluvium. |
Methodology - Canary Island fossil fishesTOP
The specimens that have been studied form part of the collection belonging to the University of Las Palmas de Gran Canaria
(Laboratory of Palaeontology) [ULPGC], with one specimen coming from the collection that belongs to El Museo Canario in Las
Palmas de Gran Canaria. We were also able to consult the Rothpletz collection in the Paläontologisches Museum München. Systematics
of the present publication for Chondrichthyes follows Cappetta (2012). The teeth terminology for Sparidae is in accordance
with Day (2002).
As the Canary marine deposits containing fossil fishes have been isotopic dated in ca 4.8 Ma, we focused the search on bio-geographic
and climatic data of the Early Pliocene, and sometimes on the more imprecise Mio-Pliocene information. We also selected one
representative locality in the Mediterrannean Sea (Lybia), two in the eastern Atlantic (Morocco and Azores), one at the Atlantic
entrance of the Mediterranean Sea and some in the Pacific coast of America just in order to show the dispersal of this fossil
fishes at that time.
Systematic palaeontologyTOP
Class CHONDRICHTHYES Huxley, 1880
Order LAMNIFORMES Berg, 1958
Family OTODONTIDAE Glickman, 1964
Carcharocles megalodon (Agassiz, 1843)
Material:
4 teeth (Fig. 6: A1, El Museo Canario); ULPGC fisches n°: f61 to f63.
|
 Fig. 6.—Chondrichthyes fossil fish teeth from Early Pliocene deposits of the Canary Islands. All from ULPGC-Pal collection, except
A from El Museo Canario collection. Scale bar: 10 mm. Carcharocles megalodon: A – Lingual view of upper right tooth. Parotodus benedeni: B - Lingual view. Cosmopolitodus hastalis: C - Labial view. Isurus oxyrinchus: D1 and D2. Lingual view. Carcharias cf. acutissima: E – First upper anterior position Lingual view. Left side of the upper jaw. Carcharhinus cf. leucas: F1 - Labial view upper lateral tooth, F2 - Anterior tooth labial view. Carcharhinus cf. priscus: G - Lingual view. Galeocerdo cf. aduncus: H1 - Lingual view. H2 - Labial view. Fig. 6.—Chondrichthyes fossil fish teeth from Early Pliocene deposits of the Canary Islands. All from ULPGC-Pal collection, except
A from El Museo Canario collection. Scale bar: 10 mm. Carcharocles megalodon: A – Lingual view of upper right tooth. Parotodus benedeni: B - Lingual view. Cosmopolitodus hastalis: C - Labial view. Isurus oxyrinchus: D1 and D2. Lingual view. Carcharias cf. acutissima: E – First upper anterior position Lingual view. Left side of the upper jaw. Carcharhinus cf. leucas: F1 - Labial view upper lateral tooth, F2 - Anterior tooth labial view. Carcharhinus cf. priscus: G - Lingual view. Galeocerdo cf. aduncus: H1 - Lingual view. H2 - Labial view.
|
|
Localities:
Barranco Seco, La Pardilla (Figs. 2– 4).
Comparisons and systematic attribution:
The teeth are large, thick and robust with a curvature towards the labial commissures in relation to the central position. The distal curvature is more marked as the position of the tooth becomes more posterior. The anterior teeth are, if at all, only very slightly curved towards the internal (lingual) face. The upper teeth are wider, slightly inclined towards the posterior edge and vertical grooves or ridges can be observed on the external (labial) face. Contrastingly, the lower teeth are narrower and more symmetrical. The crown has an equilateral triangular shape with a uniform, strongly serrated edge. The root is bilobed and coarse. All the teeth that have been found correspond to adult specimens (Gottfriend et al., 1996; Pimiento et al., 2010). Those found at La Pardilla site and kept at the Museo Canario are anterior and lateral upper teeth from the right jaw. C. megalodon is a very well known species and our specimens are identified with those studied by Menesini (1969).
Stratigraphic and geographic records:
Pan-oceanic except for the polar seas (Pimiento el al 2016). Late Miocene of Panama (Pimiento et al., 2013) Miocene and Pliocene of Morocco (Lecointre, 1952), Azores (Ávila et al., 2012), Europe (Reinecke et al., 2011), Africa (Antunes, 1978), American Atlantic and the Pacific, Ecuador (Carrillo-Briceño et al., 2014; Yabe et al., 2004).
Genus Parotodus Cappetta, 1980
Parotodus benedeni (Le Hon, 1871)
Material:
2 teeth; ULPGC n°: f71 (Fig. 6: B), f72.
Locality:
Barranco Seco (Figs. 2–4).
Comparisons and systematic attribution:
Robust, triangular crown, inclined towards the commissure. Flat labial face and markedly convex lingual face of the crown.
The crown bends towards the commissure, increasing the curvature with the lateral position of the tooth. Smooth edge. Very
thick root with two globular and separated lobules. Nutrient canal absent. Our specimens are identified with those from the
Azores shown by Ávila et al. (2012).
Stratigraphic and geographic records:
Pliocene de North America, Australia, Japan, Angola, Belgium, Italy (Cappetta, 1987), Mio-Pliocene of the Balearic Islands (Mas, 2003) and the Azores, (Ávila et al., 2012), Lower Pliocene of Huelva (Garcia et al., 2009).
Family LAMNIDAE Müller and Henle, 1838
Genus Cosmopolitodus Glikman, 1964
Cosmopolitodus hastalis (Agassiz, 1843)
Material:
41 teeth; ULPGC n°: f31 (Fig. 6: C) to f341.
Localities:
Barranco Seco, San José, Ciudad Jardín, Arenales-Chil, Bañaderos (Fig. 2–4).
Comparisons and systematic attribution:
The length from tip to base of the crown of the numerous teeth varies between 2.5 cm and 4.3 cm. The teeth are very similar to those of C. carcharias, differing in their smooth edge. The teeth are triangular, thin and compressed. The external face of the crown is concave or flat, with vertical grooves observable on rare occasions. The root is short with strongly diverging lobes that are generally
blunt and small. The teeth of the lower jaw are thicker than their upper counterparts, with a more dense and robust appearance
though their crown is narrower and rises vertically describing a slight sigmoidal curve in the anterior teeth. The external
face presents a slight concavity that is particularly noticeable in the anterior teeth. We attribute these specimens to C. hastalis given their strong resemblance to the material published by Ávila et al. (2012, Fig. 5) of Azores origin.
Stratigraphic and geographic records:
Cosmopolite Mio-Pliocene, Miocene of Peru (Klug & Kriwet, 2008), Pliocene of Belgium (Herman et al., 1974) and North America (Purdy et al., 2001). Pliocene Mediterranean (Marsili et al., 2007), Late Miocene and Early Pliocene of the Azores (Ávila et al., 2012), late Early Pliocene of Spain (García et al., 2009).
Genus Isurus Rafinesque, 1810
Isurus cf. oxyrinchus Rafinesque, 1810
Material:
4 teeth; ULPGC n°: f41 (Fig. 6: D1), f42, (Fig. 6: D2) to f44.
Locality:
Barranco Seco (Figs. 2–4).
Comparisons and systematic attribution:
The teeth display different states of preservation: none retain the root and two retain the tip of the crown. The teeth have
a narrow crown with a very sharp edge. The bulging external face is scored with longitudinal grooves and the internal face
is convex and smooth. The teeth are labially inclined and present a slight sigmoidal curvature. The lingual face resembles
that of the anterior teeth of C. hastalis, though differing in the thickness of the tooth and the bulging appearance of its external face. The narrowness of the crown
and the bulging appearance of the external face from base to tip are characteristic features of this species, along with a
slight depression at the base of the crown observable on both faces though more pronounced on the external face. Our specimens
are very similar to those shown in Marsili et al. (2007).
Stratigraphic and geographic records:
Upper Miocene and Lower Pliocene of Chile (Long, 1993) Pliocene of the south and southeast of Spain (Malaga and Catalonia) (Marsili et al., 2007). In the present day, it appears near shores with a temperate climate and tropical waters, and can migrate into warmer waters in winter (Dyldin, 2015).
Family ODONTASPIDIDAE Müller and Henle, 1839
Genus Carcharias
Rafinesque, 1810
Carcharias cf. acutissima (Agassiz, 1843)
Material:
7 incomplete teeth; ULPGC n°: f11 (Fig. 6: E), to f17
Localities:
Barranco Seco, Aljibe de la Cueva and Barranco de los Molinos (Figs. 2–5).
Comparisons and systematic attribution:
The available material is considered scarce and fragmented. The most complete specimen (Fig. 6) is a long and vertical crown 18 mm in length that is a first upper anterior tooth in the left side of the upper jaw. Even
tough dental morphology varies with the position on the jaw, the longer and vertical teeth of C. acutissima, and the most similar species C. taurus, correspond to the upper or lower anterior position (Cunningham, 2000).
For the purpose of making a roughly quantitative comparison with other similar teeth and given the longer teeth of C. acutissima, we calculated a slenderness index of the crown (width at midlength / length, on the lingual side). This index gave us a
value of ca 0.17. We compared the value of this index with that of the single specimen of the Azores (Ávila et al., 2012, Fig. 4; an upper anterior tooth) and found them to be practically identical (ca 0.16). Bauzá & Plans (1973) specimen (lám 4 Fig 28; Catalonya) is ca 0.16 as well. The corresponding slenderness index of
the most similar teeth to these, those of C. cuspitada, is ca 0.25 (Bauzá & Plans, Lám 5, Fig. 36) and C. taurus is ca 0.23 measured in a identical dental piece (Cunningham, 2000).
Stratigraphic and geographic records:
Cosmopolite Mio-Pliocene species (Cappetta, 1987). Pliocene of Morocco (Lecointre, 1952), Pliocene of Majorca (Mas, 2000), Early Pliocene of North Carolina USA (Purdy et al., 2001) and Pliocene of Ecuador (Cione et al., 2007).
Order CARCHARHINIFORMES Compagno, 1977
Family Carcharhinidae
Jordan and Evermann, 1896
Genus Carcharhinus Blainville, 1816
Carcharhinus cf. leucas (Valenciennes, 1839 in Müller and Henle, 1839–1841)
Material:
One tooth and 3 incomplete teeth; ULPGC n°: f21 (Fig. 6: F1), f22 (Fig. 6: F2), to f24.
Localities:
Barranco Seco, Ciudad Jardín, Bañaderos (Figs. 2–4).
Comparisons and systematic attribution:
Though found specimens are scarce, one of them is nearly complete (Fig. 6, F1) and corresponds to an upper anterior-lateral tooth comparable to those from the Italian Pliocene (Marsili, 2007).
The triangular shaped fragments correspond to a broad crown of small thickness. Inclination of the teeth is slightly towards
the lingual face. The lingual face is flat and the labial face slightly convex. Grooves can be seen at the base of the crown
that are more marked on the labial face. The lateral teeth have a marked curvature.
Stratigraphic and geographic records:
Miocene to Recent. They lived in warm oceans. Miocene Pacific coasts of South America (Lovejoy et al., 2006), Pliocene of North Carlolina (Purdy et al., 2001) and California (Applegate, 1978) in the USA, Angola (Antunes, 1978) and Italy (Marsili, 2007).
Carcharhinus cf. priscus (Agassiz, 1843)
Material:
One tooth; ULPGC n°: f81 (Fig. 6: G1)
Locality:
Barranco Seco (Figs. 2–4).
Comparisons and systematic attribution:
A single lateral tooth from the lower jaw with complete root. Small-sized triangular crown (0.8 cm high), smooth internal
and convex external faces. Smooth edge. Very open root. Our tooth is comparable with those from Hungary studied by Kocsis (2007).
Stratigraphic and geographic records:
Neogeno de Peru y Ecuador (Carrillo-Briceño et al., 2014), Miocene and Early Pliocene of the Mediterranean and Belgium (Menesini, 1969; Cappetta 1987). Early Pliocene of Huelva (García et al., 2011).
Genus Galeocerdo
Müller & Henle, 1837
Galeocerdo cf. aduncus Agassiz, 1843
Material:
11 teeth; ULPGC n°: f51 (Fig. 6: H1), f52 (Fig. 6: H2) to f511.
Locality:
Barranco Seco (Fig. 2–4).
Comparisons and systematic attribution:
The teeth are triangular, highly curved and in general more wide than long. The crown is markedly crenulated. A strongly serrated
talon emerges from the posterior edge, with denticles or secondary cusps whose size decreases regularly in a backwards direction.
The differences in size and inclination of the tooth become more pronounced as they are found in a more posterior position.
The anterior edge of the crown is convex and the posterior edge is strongly inclined forming an angle close to 90° with the
talon. The whole edge is heavily dentated. Crenulation is finer on the main cusp and more pronounced in the anterior region.
Denticles or secondary cusps can be observed emerging from the talon. The root is slender and presents two very open lobes
which form an angle of between 130° and 140°. It is separated from the crown by a small groove parallel to the base of the
crown. The Miocene Galeocerdo aduncus disappears in the Pliocene, giving place to G. cuvier (Menesini, 1969). However, our specimens resemble those shown in Marsili et al., (2007), while differing from those shown by Pawellek et al., (2015) particularly with respect to the angle formed by the posterior edge and the talon, which is much bigger in G. cuvier. They also differs a lot to those G. cuvier shown by Carrillo-Briceño et al., (2015; Table Fig. 3 OP Miocene and QR Early Pliocene), mainly because those are very small and long and, by contrast, the canary specimens are
nearly the same in length and in thickness. Lobule aperture at the root is wider in G. cuvier (about 160°) than in our specimens.
Stratigraphic and geographic records:
Neogene of Ecuador (Carrillo-Briceño, 2014), Miocene of Europe, America, Asia (Marsili et al., 2007), Early Pliocene of Huelva (García et al., 2011).
Class OSTEICHTHYES
Huxley, 1880
Order PERCIFORMES Johnson and Patterson, 1993
Family SPARIDAE Linné, 1758
Genus Archosargus
Gill, 1865
Archosargus cinctus (Agassiz, 1843)
Material:
593 teeth; ULPGC n°: f91 (Fig. 7: A1), f92 (Fig. 7: A2), f93 (Fig. 7: A3), f94 (Fig. 7: A4), f95 (Fig. 7: A5), f96 (Fig. 7: A6), f97 (Fig. 7: A7) to f9593.
|
 Fig. 7.—Osteichthyes fossil fish teeth from Early Pliocene deposits of Canary Islands. All from ULPGC-Pal collection. Scale bar: 10 mm. Archosargus cincta: A1 and A2 – Upper lingual hemispheroidal molariform with central depression; A3 and A4 – Upper ellipsoidal molariform of central position (between the labial and lingual series); A5 and A6 - conical teeth of the labial series A7 – 4th lower right incisiform; Labrodon pavimentatum: B1–Upper grinding face view of lower pharyngeal plate. B2 – Grinding face view of upper pharyngeal plate. B3 Lower side view of upper pharyngeal plate. B4 and B5 – Upper side view of lower pharyngeal plates. B6 – Grinding face view of upper right pharyngeal plate. Diodon scillae: C – Internal dental battery, ventral view. Fig. 7.—Osteichthyes fossil fish teeth from Early Pliocene deposits of Canary Islands. All from ULPGC-Pal collection. Scale bar: 10 mm. Archosargus cincta: A1 and A2 – Upper lingual hemispheroidal molariform with central depression; A3 and A4 – Upper ellipsoidal molariform of central position (between the labial and lingual series); A5 and A6 - conical teeth of the labial series A7 – 4th lower right incisiform; Labrodon pavimentatum: B1–Upper grinding face view of lower pharyngeal plate. B2 – Grinding face view of upper pharyngeal plate. B3 Lower side view of upper pharyngeal plate. B4 and B5 – Upper side view of lower pharyngeal plates. B6 – Grinding face view of upper right pharyngeal plate. Diodon scillae: C – Internal dental battery, ventral view.
|
|
Localities:
A few specimens appear in all the localities (Fig. 2), but the largest collection comes from the Barranco Seco (Figs. 3–4) locality with a total of 469 teeth.
Comparisons and systematic attribution:
There are three dentition types: molariform, conical and incisiform. The largest sized teeth are molariform, ranging between
42 mm and 162 mm in diameter. These are circular, hemispherical shaped and have a central depression. Some of the molariform
teeth are significantly smaller in size, ranging between 5 mm and 8 mm, while others have an oval contour and yet others a
somewhat reniform contour. A neck-like root structure with radial grooves can be observed at the base of the crown. The size
of the conical teeth is approximately half that of the molariforms. As for the incisiforms, the crown is quadrangular and
slightly broader than the root, convex and concave respectively in the external and internal faces. The concave part of the
internal face becomes wider from the base to the cutting edge where it extends along its full length.
The morphology of the Sparid fossil teeth allows us to assign them for the first time to the genus Archosargus. According to Day (2002, Fig. 13), the Sparidae which have molariform teeth belong to the genera: Sparus, Argyrops, Calamus, Archosargus, Acacthopagrus, Diplodus, Crenidens, Stenotomus, Lithognatus and Pagrus. If we eliminate the genera which possess caniniform as well as molariform teeth (Sparus, Argyrops, Calamus, Acacthopagrus, Crenidens, Stenotomus, Lithognatus and Pagrus) and the genus in which conical teeth are absent (Diplodus), the only genus that satisfies the condition of having only molariform, conical and incisiform teeth is Archosargus. In Archosargus probatocephalus (Walbaum, 1792; Worcester, 2012), a present day species of the Caribbean (Smith, 1997), the following features are observed: (a) some 120 – 130 teeth; (b) the lingual teeth are the largest with a spheroid shape
that becomes more ellipsoidal towards the anterior part; (c) the labial series is comprised of conical teeth; and (d) there
are central teeth in the maxilla of notably smaller size. There are 6 incisiforms in the upper maxilla and 8 in the lower.
From the deposits of the Early Pliocene of the Canary Islands, we have examined 382 spheroidal molariform teeth, 154 oval-shaped
molariforms, 55 conical teeth and 2 incisiform belonging to sparid fossils. The proportions and morphologies are roughly in
agreement with, on the one hand, a dentition pertaining to the genus Archosargus and, on the other with fossils attributed before the work of Day (2002) to the species Sparus cinctus and subsequently to sparid morphotypes of undetermined genus (Marsili et al., 2007). Accordingly, we consider we are justified in attributing the sparid fossils of the Canary Islands to a species of the genus
Archosargus.
Stratigraphic and geographic records:
Miocene of Morocco (Lecointre, 1952). Miocene and Pliocene of Europe and the Mediterranean, in the Atlantic from the north coast of Africa to the coast of Angola
and in the Caribbean (Menesini, 1968; Bauzà & Plans, 1973; Mas, 2000). A Lower Pliocene fossil fish found in Mediterranean Libya could be attributed to the genus Archosargus (Pawellek et al., 2015).
Family LABRIDAE Cuvier, 1816
Genus Labrodon Gervais, 1857
Labrodon pavimentatum
Gervais, 1857
Material:
63 pharyngeal plates; ULPGC n°: f101 (Fig. 7: B1), f102 (Fig. 7: B2), f103 (Fig. 7: B3), f104 (Fig. 7: B4), f105 (Fig. 7: B5 and B6) to f1063.
Localities:
Barranco Seco, Ciudad Jardín (Figs. 2–4).
Comparisons and systematic attribution:
Among the numerous pharyngeal plates found in various states of conservation we have chosen for representation purposes: three
lower pharyngeal plates, and one right and one left upper pharyngeal plates (Fig. 7, B). These plates were described in detail by Cocchi (1864), who based his species Pharyngodopilus africanus on material from the same Barranco Seco site on the island of Gran Canaria. In some of our specimens, there also appears
a sigmoidal shape similar to that of Pharyngodopilus crassus. The Barranco Seco pharyngeal plates are very similar, if not identical, to L. multide, according to Sacco (1916), and to L. pavimentatum, according to Simonelli (1889). Moreover, according to Sacco (1916), Pharyngodopilus crassus is synonymous with L. pavimentatum.
Stratigraphic and geographic records:
Very abundant in Miocene deposits of the Mediterranean coasts (Cocchi, 1864). Miocene of France and Portugal (Sauvage, 1875). Miocene of Costa Rica (Laurito et al., 2014). Pliocene of Morocco (Lecointre, 1952).
Order TETRAODONTIFORMES Regan, 1929
Family DIODONTIDAE Bibron in Duméril, 1855
Genus Diodon Linné 1758
Diodon scillae
Agassiz, 1843
Material:
2 complete dental plates and one fragment; (Fig. 1, Paläontologisches Museum München) and; ULPGC n°: f111 (Fig. 7: C1) to f113.
Localities:
Barranco de Guiniguada, Barranco Seco (Fig. 2–4).
Comparisons and systematic attribution:
Complete upper dental plate, approximately 18 mm wide. Bilobed structure with heart-shaped contour and triangular-shaped lobes
comprised of 14–16 fused blades or platelets. The uppermost pair of platelets are very thin and narrow whereas the two at
the base are wider and ellipsoidal or trapezoidal in shape. The specimens are identical to that described by Rothpletz & Simonelli (1890) from Gran Canaria island, which is currently part of the PMM collection [Paläontologisches Museum München] (Fig. 1) and very similar to the specimen from the Miocene of Cuba in shape and dimensions. D. scillae is a senior synonym of D. sigma (Iturralde-Vinent & Case, 1998).
Stratigraphic and geographic records:
Miocene of France (Leriche, 1957), Italy (Menesini, 1969), Miocene of Cuba (Iturralde-Vinent & Case, 1998).
DiscussionTOP
The presence of pillow lavas in relation to the deposits (Meco et al., 2007, 2015) has allowed a dating (ca 4.8 Ma) for Canary fossil fishes, which - from the 19th through to the 21st century - have gone from being attributed to
the “Miocene′ (Fig. 1), “Mio-Pliocene′ or “Neogene′ to finally being assigned to the first half of the Early Pliocene.
The age of the Canary deposits allows a reconstruction of the palaeoceanographic scenario in which the fossil fishes considered
in this paper lived. A means of communication was open at the time between the Atlantic and Pacific Oceans via the CAS (Erkens, 2015), while the Gibraltar Seaway was again connecting the Atlantic Ocean with the Mediterranean Sea (Garcia-Castellanos et al., 2009). In addition to these circumstances, we need to consider the capacity of some of these fishes to travel large distances.
About 5 to 4 Ma ago, in the Early Pliocene, the earth had a warm, temperate climate (Fedorov et al., 2013). The marine waters of the Canary Islands between 4.8 and 4.2 Ma (Meco et al., 2015, 2016), as well as those of the Azores (Santa Maria Island), were subtropical to warm-temperate seas (Ávila et al., 2012). The presence in the Mediterranean (Lybia) of the genera Galeocerdo, Carcharhinus, Archosargus (?) and Diodon indicates a tropical to equatorial climate (Pawellek et al., 2012).
Among the Chondrichthyan fishes studied we find Carcharocles megalodon and Isurus hastalis, the largest marine predators of the Pliocene (Randall, 1987; Gottfried et al., 1996). It is thought that these were trophic migrators with a pan-oceanic distribution who sought out stable food sources. All
of the above lived in all seas during the Early Pliocene. Notable among the Osteichthyes is the presence of the genus Archosargus. The few present day species of this genus live mainly in the western Atlantic, the Caribbean and the Galapagos islands (Smith, 1997). Their presence in the eastern Atlantic and the Mediterranean would be consonant with the palaeogeographic conditions described
above (Meco et al., 2015) and related to the CAS. One possible Archosargus has been found in the Early Pliocene of Libya (Pawellek et al., 2012). Diodon scillae has been found in the Miocene of Cuba and the Mediterranean (Iturralde-Vinent & Case, 1998).
The coincidental circumstances of an open CAS and Gibraltar Seaway, along with the capacity of the Chondrichthyes to travel
long distances, highlight the tropicopolitan nature of Canary fossil fishes in the Early Pliocene. This is a factor to be
considered in the explanation of the presence of numerous Indo-Pacific sharks in the Mediterranean (Pawellek et al., 2012).
The teeth of the genus Galeocerdo found in Gran Canaria Island are comparable to G. aduncus which is a Miocene species (Marsili et al., 2007). For Cigala-Fulgosi & Mori (1979), the main difference between this fossil species and the present-day species (G. cuvier) lies in the greater robustness of the teeth of G. cuvier. Nonetheless, many teeth attributed to G. aduncus due to their stratigraphic origin cannot be distinguished from those of young, small or immature specimens of G. cuvier. According to Cigala-Fulgosi & Mori (1979), the transition or evolution from G. aduncus to G. cuvieri was very rapid and possibly coincided with the start of the Early Pliocene (5.33 Ma); both are present in deposits of the
Late Miocene whereas only G. cuvier has been found in Early Pliocene deposits. Consequently, the presence of G. aduncus in the Early Pliocene of Gran Canaria (ca 4.80 Ma) could be added to those from the Early Pliocene of Huelva (SW Spain) (Garcia et al., 2011).
ConclusionTOP
A total of 8 taxa of Chondrichthyes and 3 of Osteichthyes have been identified and classified: Carcharocles megalodon, Parotodus benedeni, Cosmopolitodus hastalis, Isurus oxyrinchus, Carcharias cf. acutissima, Carcharhinus cf. leucas, Carcharhinus cf. priscus, Galeocerdo cf. aduncus, Archosargus cinctus, Labrodon pavimentatum and Diodon scillae.
Of seven fossil shark species found in the Azores and eight in the Canary Islands, six or seven are common to both archipelagos
(Carcharocles megalodon, Parotodus benedeni, Cosmopolitodus hastalis, Isurus oxyrinchus Carcharias cf. acutissima, Carcharhinus cf. leucas, and perhaps Galeocerdo cf. aduncus = Carcharhinus egertoni?) (Fig. 8).
|
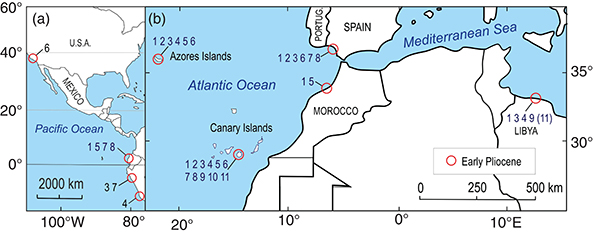 Fig. 8.—Maps of selected localities with palaeogeographic significance: (a) on the western Pacific coasts; (b) on the eastern Atlantic
and Mediterranean coasts. Numbers refers to the fossil species of the present paper: 1 Carcharocles megalodon, 2 Parotodus benedeni, 3 Cosmopolitodus hastalis, 4 Isurus oxyrinchus, 5 Carcharias cf. acutissima, 6 Carcharinus cf. leucas, 7 Carcharinus cf. priscus, 8 Galeocerdo cf. aduncus, 9 Diodon scillae, 10 Labrodon pavimentatum, 11 Archosargus cinctus. Fig. 8.—Maps of selected localities with palaeogeographic significance: (a) on the western Pacific coasts; (b) on the eastern Atlantic
and Mediterranean coasts. Numbers refers to the fossil species of the present paper: 1 Carcharocles megalodon, 2 Parotodus benedeni, 3 Cosmopolitodus hastalis, 4 Isurus oxyrinchus, 5 Carcharias cf. acutissima, 6 Carcharinus cf. leucas, 7 Carcharinus cf. priscus, 8 Galeocerdo cf. aduncus, 9 Diodon scillae, 10 Labrodon pavimentatum, 11 Archosargus cinctus.
|
|
In addition, we have also identified among the Osteichthyan fishes from the Canary Islands, thanks to the numerous teeth and
dental fragments that have been found the tropical genera Archosargus and Diodon.
The fossil species found in the Early Pliocene of Libya that have also been found in Gran Canaria are: Carcharocles megalodon, Cosmopolitodus hastalis, Isurus oxyrinchus, genera Archosargus cinctus and Diodon scillae.
Carcharocles megalodon, Cosmopolitodus hastalis, Isurus cf. oxyrinchus, Carcharias cf. acutissima, Carcharhinus cf. leucas, Carcharhinus cf. priscus, Galeocerdo cf. aduncus were also present in the Pliocene Pacific coasts of South America. Consequently, their preference for warm waters
suggests an open CAS between the Atlantic and Pacific oceans at that time.
Galeocerdo adunctus from the Canary Early Pliocene should to be added to the reference from the same epoch in Huelva (SW mainland Spain).
ACKNOWLEDGEMENTSTOP
This work was sponsored by the Spanish Ministry of the Environment and the University of Las Palmas de Gran Canaria (CN-62/03-02139).
We thank the curators of the Paläontologisches Museum München, Bayerische Staatssammlung für Paläontologie und Geologie and
of El Museo Canario. We also thank the comments from the Editor and the suggestions from the reviewers.
ReferencesTOP
| ○ |
Agassiz, L. (1833–1843). Recherches sur les poissons fossiles. Petitpierre, Neuchâtel. 1420 p. http://dx.doi.org/10.5962/bhl.title.4275
|
| ○ |
Anguita Virella, F. & Ramírez del Pozo, J. (1974). La datación micropaleontológica de la Terraza de Las Palmas (Gran Canaria).
Estudios Geológicos, 30: 185–188.
|
| ○ |
Antunes, M.T. (1978). Faunes ichthyologiques du Néogène supérieur d’Angola, leur âge, remarques sur le Pliocène marin en Afrique
australe. Ciências da Terra (UNL), 4: 59–90. http://hdl.handle.net/10362/1501
|
| ○ |
Applegate, S.P. (1978). Phyletic studies, parte I. Tiger Shark. Revista - Instituto de Geología. Universidad Nacional Autonoma
de México, 2 (1): 55–64.
|
| ○ |
Ávila, S.P.; Ramalho, R. & Vullo, R. (2012). Systematic, palaeoecology and palaeobiogeography of the Neogene fossil sharks
from the Azores (Northeast Atlantic). Annales de Paléontologie, 98 (3): 167–189. http://dx.doi.org/10.1016/j.annpal.2012.04.001
|
| ○ |
Bauzà, J. & Plans, J. (1973). Contribución al conocimiento de la fauna ictiológica del Neógeno catalano-balear. Boletín de
la Sociedad de Historia Natural de Baleares, 18: 72–131.
|
| ○ |
Berg, L.S. (1958). System der regenten und fossilen Fischartigen und Fische. Deutsch Verlag Wiss, Berlin, 310 p.
|
| ○ |
Betancort, J.F.; Lomoschitz, A. & Meco, J. (2014). Mio-Pliocene crustaceans from the Canary Islands, Spain. Rivista Italiana
di Paleontologia e Stratigrafia, 120 (3): 337–349. http://dx.doi.org/10.13130/2039-4942/6076
|
| ○ |
Blainville, H.D.D. (1816). Prodrome d’une nouvelle distribution systematique de regne animal. Bulletin des Sciences, par la
Société Philomatique. Paris, 8: 113–124. http://www.biodiversitylibrary.org/part/18415
|
| ○ |
Cappetta, H. (1980). Modification du statut générique de quelques espèces de sélaciens crétacés et tertiaires. Palaeovertebrata,
10 (1): 29–42.
|
| ○ |
Cappetta, H. (1987). Chondrichthyes II: Mesozoic and Cenozoic Elasmobranchii. In: Schultze, H.P. (Ed.). Handbook of Paleoichthyology.
Gustav Fischer Verlag, Stuttgart, 1–193.
|
| ○ |
Capetta, H. (2012). Chondrichthyes: Mesozoic and Cenozoic Elasmobrachii: Teeth. In: Schultze, H.P. (Ed.). Handbook of Paleoichthyology.
Gustav Fischer Verlag, Stuttgart. 512 p.
|
| ○ |
Carracedo, J.C.; Pérez Torrado, F.J.; Ancochea, E.; Meco, J.; Hernán, F.; Cubas, C.R.; Casillas, R.; Rodríguez Badiola, E.
& Ahijado, A. (2002). Cenozoic volcanism II: the Canary Islands. In: Gibbons, W. & Moreno, T. (Eds.). The Geology of Spain.
Geological Society, London, 439–472. http://dx.doi.org/10.1144/GOSPP.18
|
| ○ |
Carrillo-Briceño, J.D.; Aguilera, O.A. & Rodríguez, F. (2014). Fossil Chondrichthyes from the central Eastern Pacific Ocean
and their paleoceanographic significance. Journal of South American Earth Sciences, 51: 76–90. http://dx.doi.org/10.1016/j.jsames.2014.01.001
|
| ○ |
Carrrillo-Briceño, J.D.; Maxwell, E.; Aguilera, O.A.; Sánchez, R. & Sánchez-Villagra, M.R. (2015). Sawfisches and Other Elasmobranch
Assemblages from the Mio-Pliocene of the South Caribbean (Urumaco Sequence, Northwestern Venezuela). PLoS ONE, 10 (10): e0139230.
http://dx.doi.org/10.1371/journal.pone.0139230
|
| ○ |
Cigala-Fulgosi, F. & Mori, D. (1979). Osservazioni tassonomiche sul genere Galeocerdo (Selachii, Carcharhinidae) con particolare
riferimento a Galeocerdo cuvieri (Péron & Lesueur) nel Pliocene del Mediterraneo. Bollettino della Società Paleontologica
Italiana, 18: 117–132.
|
| ○ |
Cione, A.L.; Mennuccci, J.A.; Santalucita, F. & Hospitalache, C.A. (2007). Local extintion of sharks of genus Carcharias Rafinesque,
1810 (Elasmobranchii, Odontaspididae) in the easteren Pacific Ocean. Revista geológica de Chile, 34 (1): 139–145.
|
| ○ |
Cocchi, I. (1864). Monographia dei Pharyngodopilidae. Nuova famiglia di pesci Labroidi. Cellini, Firenze, 88 p.
|
| ○ |
Compagno, L.J.V. (1977). Phyletic relationships of living sharks and rays. American Zoologist, 17: 303–322. http://dx.doi.org/10.1093/icb/17.2.303
|
| ○ |
Cunningham, S.B. (2000). A comparison of isolated teeth of early Eocene Striatolamia macrota (Chondrichthyes, Lamniformes),
with those of a Recent sand shark, Carcharias taurus. Tertiary Research, 20: 17–31.
|
| ○ |
Cuvier, G.L.C.E.D. (1816). Le Règne Animal Distribué D’après son Organisation Pour Servir de Base à L’histoire Naturelle des
Animaux et D’introduction à L’anatomie Comparée. T. II, Les reptiles, les Poissons, les Mollusques et les Annélides. Deterville,
Paris, 532 p. http://biodiversitylibrary.org/page/1848835
|
| ○ |
Day, J.J. (2002). Phylogenetic relationships of the Sparidae (Teleostei: Percoidei) and implications for convergent trophic
evolution. Biological Journal of the Linnean Society, 76: 269–301. http://dx.doi.org/10.1111/j.1095-8312.2002.tb02088.x
|
| ○ |
Duméril, A. (1855). Note sur un travail inédit de Bibron relatif aux poissons plectognathes gymnodontes (diodons et tétrodons).
Revue et Magasins de Zoologie, série 2, 8: 274–282.
|
| ○ |
Dyldin, Y.V. (2015). Annotated checklist of the sharks, batoids and chimaeras (Chondrichthyes: Elasmobranchii, Holocephali)
from waters of Russia and adjacent areas. Publications of the Seto Marine Biological Laboratory, 43: 40–91. http://hdl.handle.net/2433/197957
|
| ○ |
Erkens, R.H.J. (2015). The less-splendid isolation of the South America continent. Frontiers of Biogeography, 7 (3): 89–90.
http://eprints.cdlib.org/uc/item/46g0b2tj
|
| ○ |
Fedorov, A.V.; Brierley, C.M.; Lawrence, K.T.; Liu, Z.; Dekens, P.S. & Ravelo, A.C. (2013). Patters and mechanisms of early
Pliocene warmth. Nature, 496: 43–49. http://dx.doi.org/10.1038/nature12003
|
| ○ |
Fritsch, K. von (1867). Reisebilder von den Canarischen Inseln. Petermann’s geographischen Mittheilungen, 22: 1–43.
|
| ○ |
García, E.X.M.; Telles-Antunes, M.; Cáceres-Balbino, A.; Ruiz-Muñoz, F. & Civis-Llovera, J. (2009). Los tiburones Lamniformes
(Chondrichthyes, Galeomorphii) del Plioceno inferior de la Formación Arenas de Huelva, suroeste de la cuenca del Guadalquivir,
España. Revista Mexicana de ciencias geológicas, 26 (3): 674–686.
|
| ○ |
García-Castellanos, D.; Estrada, F.; Jiménez-Munt, C.; Fernàndez, M.; Vergés, J. & De Vicente, R. (2009). Catastrophic flood
of the Mediterranean after Messinian salinity crisis. Nature, 462: 778–781. http://dx.doi.org/10.1038/nature08555
|
| ○ |
Gervais, P. (1857). Sur un poisson labroide fossile dans les sables marins de Montpellier (Labrodon pavimentatum). Mémoires
de la Académie des Sciences de Montpellier, 3, 515 p.
|
| ○ |
Gill, T.N. (1865). Synopsis of the pleuronectoids of California and Northwestern America. Proceedings of the Academy of Natural
Sciences of Philadelphia, 16 (4): 194–198. http://www.jstor.org/stable/4623938
|
| ○ |
Gottfried, M.D.; Compagno, L.J.V. & Bowman, S.C. (1996). Chapter 7: Size and skeletal anatomy of the Giant Megatooth shark
Carcharodon megalodon. In: Klimley, A.P. & Ainley, D.G. (Eds.). Great White Sharks: the Biology of Carcharodon carcharias.
Academic Press, San Diego, 55–66. http://dx.doi.org/10.1016/B978-012415031-7/50008-2
|
| ○ |
Gradstein, F.M.; Ogg, J.G.; Smith, A.G.; Agterberg, F.P.; Bleeker, W.; Cooper, R.A.; Davydov, V.; Gibbard, P.; Hinnov, L.;
House, M.R.; Lourens, L.; Luterbacher, H.P.; McArthur, J.; Melchin, M.J.; Robb, L.J.; Shergold, J.; Villeneuve, M.; Wardlaw,
B.R.; Ali, J.; Brinkhuis, H.; Hilgen, F.J.; Hooker, J.; Howarth, R.J.; Knoll, A.H.; Laskar, J.; Monechi, S.; Plumb, K.A.;
Powell, J.; Raffi, I.; Röhl, U.; Sanfilippo, A.; Schmitz, B.; Shackleton, N.; Shields, G.A.; Strauss, H.; Van Dam, J.; van
Kolfschoten, T.; Veizer, J. & Wilson, D. (2004). A geologic time scale 2004. Cambridge University Press, Cambridge. 589 p.
|
| ○ |
Herman, J.; Crochard, M. & Girardot, M. (1974). Quelques restes de Sélaciens récoltés dans les sables du Kattendijk à Kallo.
Bulletin de la Société Belge de Géologie, 83 (1): 15–31.
|
| ○ |
Huxley, T.H. (1880). On the applications of the laws of evolution to the arrangement of the vertebrata and more particularly
of the Mammalia. Proceedings of the Zoological Society of London, 43: 649–661. http://biodiversitylibrary.org/page/28522869
|
| ○ |
Iturralde-Vinent, M. & Case, G.R. (1998). First report of the fossil fish Diodon (Family Diodontidae) from the Miocene of
Cuba. Revista de la Sociedad Mexicana de Paleontología, 8: 123–126.
|
| ○ |
Johnson, G.D. & Patterson, C. (1993). Percomorph phylogeny: A survey of acanthomorphs and a new proposal. Bulletin of Marine
Science, 52 (1): 554–626.
|
| ○ |
Jordan, D.S. & Evermann, B.W. (1896). The fishes of North and Middle America. Bulletin of the United States National Museum,
47: 1–1240.
|
| ○ |
Klug, S. & Kriwet, J. (2008). Skeletal remains of the Miocene lamniform shark, Cosmopolitodus hastalis, from Peru. Erlanger
geologische Abhandlungen, Sonderband, 6: 99.
|
| ○ |
Kocsis, L. (2007). Central Paratethyan shark fauna (Ipolytarnóc, Hungary). Geologica Carpathica, 58 (1): 27–40.
|
| ○ |
Laurito, C.A.; Calvo, C.; Valerio, A.L.; Calvo, A. & Chacón, R. (2014). Ictiofauna del mioceno inferior de la localidad de
Pacuare de Tres Equis, formación río Banano, provincia de Cartago, Costa Rica, y descripción de un nuevo género y una nueva
especie de scaridae. Revista Geológica de América Central, 50: 153–192. http://dx.doi.org/10.15517/rgac.v0i50.15121
|
| ○ |
Lecointre, G. (1952). Recherches sur le Néogène et le Quaternaire marin de la Côte Atlantique du Maroc. Paléontologie. Notes
et Mémoires, Service géologique du Maroc, 99: 1–172.
|
| ○ |
Le Hon, H. (1871). Préliminaires d’une Mémoire sur les Poissons tertiaires de Belgique. H. Merzbach, Bruxelles, 15 p.
|
| ○ |
Leriche, M.E. (1957). Les poissons néogènes de la Bretagne de l’Anjou et de la Touraine. Mémoires de la Société Géologique
de France, 81: 1–64.
|
| ○ |
Linnæus, C. (Linné) (1758). Systema naturæ per regna tria naturæ, secundum classes, ordines, genera, species, cum characteribus,
differentiis, synonymis, locis. Tomus I. Editio decima, reformata. Holmiæ, Stockholm. 532 p.
|
| ○ |
Lomoschitz, A.; Betancort, J.F.; Cilleros, A. & Meco, J. (2011). Sedimentology of a Pliocene volcano-sedimentary sequence
on the NE coast of Gran Canaria (Canary Islands, Spain) and its paleogeographic significance. 28th IAS Meeting on Sedimentology,
Abstracts Volume, Zaragoza, 165.
|
| ○ |
Long, D.J. (1993). Late Miocene and Early Pliocene fish assemblages from the north central coast of Chile. Tertiary Research
(United Kingdom), 14: 117–126.
|
| ○ |
Lovejoy, N.R. & Albert, J.S. (2006). Miocene marine incursions and marine/freshwater transitions: Evidence from Neotropical
fishes. Journal of South American Earth Sciences, 21 (1–2): 5–13. http://dx.doi.org/10.1016/j.jsames.2005.07.009
|
| ○ |
Lyell, C. (1865). Elements of Geology, 6th ed., John Murray, London, 196 p.
|
| ○ |
Marsili, S. (2007). Revision of the teeth of the genus Carcharhinus (Elasmobranchii; Carcharhinidae) from the Pliocene of
Tuscany, Italy. Rivista Italiana di Paleontologia e Stratigrafia, 113: 79–95. http://dx.doi.org/10.13130/2039-4942/6360
|
| ○ |
Marsili, S.; Carnevale, G.; Danese, E.; Bianucci, G. & Landini, W. (2007). Early Miocene vertebrates from Montagna della Maiella,
Italy. Annales de Paléontologie, 93 (1): 27–66. http://dx.doi.org/10.1016/j.annpal.2007.01.001
|
| ○ |
Martin, K. (1887). Palaeontologische Ergebnisse von Tiefbohrungen auf Java, nebst allgemeineren Studien ueber das Tertiaer
von Java, Timor und einiger anderer Inseln. Leiden, 380 p.
|
| ○ |
Mas, G. (2000). Ictiofauna del Pliocè mitjà-superior de la conca sedimentària de Palma (Illes Balears, Mediterrània Occidental).
Implicacions paleoambientals. Bolletí de la Societat d’Història Natural de les Balears, 43: 39–61.
|
| ○ |
Mas, G. (2003). Presència de Parotodus benedeni (Le Hon, 1871) (Pisces: Chondrichthyes: Otodontidae) al Neogen de Mallorca
i Menorca (Illes Balears, Mediterrània occidental). Consideracions taxonòmiques i paleoambientals. Bolletí de la Societat
d’Història Natural de les Balears, 46: 85–90.
|
| ○ |
Meco, J. (1977). Paleontología de Canarias I: Los Strombus neógenos y cuaternarios del Atlántico euroafricano: taxonomía,
bioestratigrafía y paleoecología. Cabildo Insular de Gran Canaria, Las Palmas de Gran Canaria. 142 p.
|
| ○ |
Meco, J. (1981). Neogastrópodos fósiles de las Canarias orientales. Anuario de Estudios Atlánticos, 27: 601–615.
|
| ○ |
Meco, J. (1982). Los Bivalvos fósiles de las Canarias orientales. Anuario de Estudios Atlánticos, 29: 579–595.
|
| ○ |
Meco, J. & Stearns, C.E. (1981). Emergent littoral deposits in the Eastern Canary Islands. Quaternary Research, 15 (2): 199–208.
http://dx.doi.org/10.1016/0033-5894(81)90104-6 |
| ○ |
Meco, J.; Scaillet, S.; Guillou, H.; Lomoschitz, A.; Carracedo, J.C.; Ballester, J.; Betancort, J.F. & Cilleros, A. (2007). Evidence for long-term uplift on the Canary Islands from emergent Mio-Pliocene littoral deposits. Global and Planetary Change, 57 (3–4): 222–234. http://dx.doi.org/10.1016/j.gloplacha.2006.11.040
|
| ○ |
Meco, J.; Koppers, A.A.P.; Miggins, D.P.; Lomoschitz, A. & Betancort, J.F. (2015). The Canary Record of the Evolution of the North Atlantic Pliocene: New 40Ar/39Ar Ages and Some Notable Palaeontological Evidence. Palaeogeography, Palaeoclimatology, Palaeoecology, 435: 53–69. http://dx.doi.org/10.1016/j.palaeo.2015.05.027
|
| ○ |
Meco, J.; Lomoschitz, A. & Betancort, J.F., (2016). Early Pliocene tracer of North Atlantic and South Pacific sea surface currents: Janthina typica (Bronn, 1860) (Mollusca Gastropoda). Revista Mexicana de Ciencias Geológicas, 33 (2): 192–197.
|
| ○ |
Menesini, E. (1969). Ittiodontoliti miocenici di Terra d’Otranto (Puglia). Palaeontolographia Italica, 65: 1–61.
|
| ○ |
Montes, C.; Bayona, G.A.; Cardona, A.A.; Bush, D.M.; Silva, C.A.; Morón, S.E.; Hoyos, N.; Ramírez, D.A.; Jaramillo, C.A. & Valencia, V. (2012). Arc-continent collision and orocline formation: closing of the Central American seaway. Journal of Geophysical Research, 117: B04105. http://dx.doi.org/10.1029/2011JB008959
|
| ○ |
Müller, J. & Henle, J. (1837). Gattungen der Haifische und Rochen nach einer von ihm mit Hrn. Henle unternommenen gemeinschaftlichen Arbeit tiber die Naturgeschichte der Knorpelfische. Akademie der Wissenschaften Berlin, 2: 111–118.
|
| ○ |
Müller, J. & Henle, F.G.J. (1838). On the generic characters of the cartilagi- nous fishes, with descriptions of the new genera. Magazine of Natural History, 2: 36.
|
| ○ |
Müller, J. & Henle, F.G.J. (1839). Systematische Beschreibung der Plagiostomen. Veit, Berlin, 2: 73.
|
| ○ |
Pawellek, T.; Adnet, S.; Cappetta, H.; Metais, E.; Salem, M.; Brunet, M. & Jaeger, J.J. (2012). Discovery of an earliest Pliocene relic tropical fish fauna in a newly detected Cliff section (Sabratah Basin, NW Libya). Neues Jahrbuch fur Geologie und Paläeontologie Abhandlungen, 266: 93–114. http://dx.doi.org/10.1127/0077-7749/2012/0272
|
| ○ |
Pimiento, C.; Ehret, D.J.; Hendy, A.J.W.; MacFadden, B.J.; Bruce, J. & Hubbel, G. (2010). Ancient Nursey Area for the Extinct
Giant Shark Megalodon from the Miocene of Panama. PLoS One 5 (5): e10552. http://dx.doi.org/10.1371/journal.pone.0010552
|
| ○ |
Pimiento, C.; González-Barba, G.; Ehret, D.J.; Hendy, A.J.W.; MacFadden, B.J. & Jaramillo. C. (2013). Sharks and Rays (Chondrichthyes,
Elasmobranchii) from the Late Miocene Gatun Formation of Panama. Journal of Paleontology, 87: 755–774. http://dx.doi.org/10.1666/12-117
|
| ○ |
Pimiento, C.; MacFadden, B.J., Clemens, C.F.; Varela, S.; Jaramillo, C.; Velez-Juarbe, J. & Silliman, B.R. (2016). Geographical
distribution patterns of Carcharocles megalodon over time rveal clues about extinction mechanisms. Journal of Biogeography,
43 (8): 1645–1655. http://dx.doi.org/10.1111/jbi.12754
|
| ○ |
Purdy, R.; Schneider, V.P.; Applegate, S.P.; McLellan, J.H.; Meyer, R.L., & Slaughter, B.H. (2001). The Neogene sharks, rays,
and bony fishes from Lee Creek Mine, Aurora, North Carolina. In: Ray, C.E. & Bohaska, D.J. (Eds.). Geology and Paleontology
of the Lee Creek Mine, North Carolina. Smithsonian Contributions to Paleobiology, 90. Smithsonian Institution Press, Washington
D.C., 71–202.
|
| ○ |
Rafinesque, C.S. (1810). Caratteri di alcuini nuovi generi e nuove spece di animali e piante della Sicilia. Parte 1. Stampe
di Sanfilippo, Palermo, 105 p. http://dx.doi.org/10.5962/bhl.title.104418
|
| ○ |
Randall, J.E. (1987). Refutation of lengths of 11.3, 9.0, and 6.4 m attributed to the white shark, Carcharodon carcharias.
California Fish and Game, 73: 169–174.
|
| ○ |
Regan, C.T. (1929). Fishes. Encyclopaedia Brittanica, 9. 14th ed., Encyclopaedia Brittanica Co., London, 305–329.
|
| ○ |
Reinecke, T.; Louwye, S.; Havekost, U. & Moths, H. (2011). The elasmobranch fauna of the late Burdigalian, Miocene, at Werde-Uesen,
Lower Saxony, Germany, and its relationships with Early Miocene faunas in the North Atlantic, Central Parathethys and Mediterranean.
Palaeontos, 20: 1–170.
|
| ○ |
Rothpletz, A. & Simonelli, V. (1890). Die marinen Ablagerungen auf Gran Canaria. Zeitschrift der Deutschen geologischen Gesellschaft,
42 (4): 677–736.
|
| ○ |
Sauvage, H-E. (1875). Note sur le genre Nummopalatus et sur les espèces de ce genre trouvées dans les terrains tertiaires
de la France. Bulletin de la Société géologique de France, 3: 613–642.
|
| ○ |
Sacco, F. (1916). Apparati dentali di “Labrodon’’ e di “Chrysophrys’’ del Pliocene italiano. Reale Academia delle Science
di Torino, 60: 144–149.
|
| ○ |
Schmidt, D.N. (2007). The closure history of the Central American seaway: evidence from isotopes and fossils models and molecules.
In: Williams, M.; Wood, A.M.; Gregory, J.F. & Schmidt, D.N. (Eds.). Deep time perspectives on climate change: marrying the
signal from computer models and biological proxies. Geological Society of London, London, 427–442.
|
| ○ |
Sendino, C. & Taylor, P.D. (2014). Pliocene bryozoans from Gran Canaria. Studi Trentini di Scienze Naturali, 94: 229–239.
|
| ○ |
Simonelli, V. (1889). Terreni e fossili di Pianosa nel mar Tirreno. Bolletino del Reale Comitato Geolologico d’Italia, 10:
193–237.
|
| ○ |
Smith, C.L. (1997). National Audubon Society field guide to tropical marine fishes of the Caribbean, the Gulf of Mexico, Florida,
the Bahamas, and Bermuda. Alfred A. Knopf, Inc., New York. 720 p.
|
| ○ |
Walbaum, J.J. (1792). Petri Artedi renovati. Part 3. Petri Artedi sueci genera Piscium in quibus systema totum ichthyologiae
proponitur cum classibus, ordinibus, generum characteribus, specierum diffentiis, observationibus plumiris. Redactis Speciebus
2. Ichthyologiae, part III. Grypeswaldiae, 723 p.
|
| ○ |
Walker, J.D. & Geissman, J.W. (2009). 2009 Geologic Time scale. Geological Society of America, http://dx.doi.org/10.1130/2009.CTS004R2C
|
| ○ |
Worcester, C.E. (2012). Phenotypic Plasticity of Oral Jaw Dentition in Archosargus Probatocephalus. PhD Thesis, Western Kentucky
University, 38 p. http://digitalcommons.wku.edu/theses/1215
|
| ○ |
Yabe, H.; Goto, M. & Kaneko, N. (2004). Age of Carcharocles megalodon (Lamniformes: Otodontidae): A review of the stratigraphic
records (text in Japanese with abstract in English). Fossils (Tokyo), 75: 7–15.
|