Trace fossils and dubiofossils from the Ediacaran and Cambrian of the Alcudia Anticline, Spain
Pistas fósiles y dubiofósiles del Ediacárico y Cámbrico del Anticlinal de Alcudia, España
J. Simón
Dpto. Biología y Geología. IES Peñalba. Molinos de Viento s/n, Moral de Calatrava, Ciudad Real, Spain. Email: jsimon@edu.jccm.es. ORCID ID: http://orcid.org/0000-0002-0064-5672
| |
ABSTRACT
The fossil record of the Ediacaran—Cambrian transition in Spain provides potentially useful information to understand this critical episode in the history of life on Earth. In the present work, new findings of trace fossils and dubiofossils are described from the upper Ediacaran and lowermost Cambrian of the Alcudian unit in the Alcudia Anticline, at the southern margin of the Central Iberian Zone (valle de Alcudia, Iberian Massif, Spain). Two new trace fossil sites are described from the Upper Alcudian subunit and assigned to the Fortunian stage (lower Cambrian, Terreneuvian series) and to the regional Lower Corduban substage. They contain examples of arthropod-like scratches (Monomorphichnus lineatus), small bilobed trails (similar to Archaeonassa), an inclined burrow with spreite (may be a teichichnid), and long unbranched burrows with very fine transversal structure (probably the ventral surface of Psammichnites). Torrowangea aff. rosei and possible body fossils of late Ediacaran age are described from the Lower Alcudian subunit, including a disc resembling a frond holdfast and a diamond-shaped complex structure with no known equivalents.
Keywords: Iberian Massif; Lower Cambrian; Precambrian-Cambrian transition; Terreneuvian.
|
| |
RESUMEN
El registro fósil de la transición Ediacárico—Cámbrico en España proporciona información potencialmente útil para entender este episodio crítico de la historia de la vida en La Tierra. El presente trabajo describe nuevos hallazgos de icnofósiles y dubiofósiles del Ediacárico superior y Cámbrico basal en la unidad Alcudiense del Anticlinal de Alcudia, hacia el margen meridional de la Zona Centroibérica (valle de Alcudia, Macizo Ibérico, España). Dos localidades con pistas fósiles son descritas para la subunidad Alcudiense Superior, y asignadas al piso Fortuniense (Cámbrico inferior, serie Terreneuviense) y al subpiso regional Cordubiense Inferior. Contienen ejemplos de marcas de artrópodos u organismos similares (Monomorphichnus lineatus) y pistas bilobadas simples (semejantes a Archaeonassa), una madriguera con spreite inclinada (quizá un teichíchnido) y madrigueras largas, sin ramificar, con estructura transversal muy fina (probablemente la superficie ventral de Psammichnites). Se describen Torrowangea aff. rosei y posibles fósiles corporales de edad ediacárica tardía en la subunidad Alcudiense Inferior, incluyendo un disco semejante al anclaje de un fronde, y una estructura compleja, con forma de diamante, sin equivalentes conocidos.
Palabras clave: Macizo Ibérico; Cámbrico inferior; Terrenouviense; Transición Precámbrico-Cámbrico.
|
IntroductionTOP
The transition from the Precambrian biosphere, dominated by microorganisms, to the Phanerozoic world, where animals are a keystone group from a biogeochemical and evolutionary perspective, is arguably the deepest ecological change in the history of life on Earth (Crimes, 1992; Fedonkin et al., 2007; Seilacher, 2007). This revolution was mainly concentrated in the Ediacaran—Cambrian transition, whose fossil record in central Spain is not very abundant but quite diverse, containing stromatolites, trace fossils, soft-bodied organisms (vendotaenids, sabelliditids, acritarchs, possible Beltanelliformis), and early skeletal fossils (Cloudina, Sinotubulites, Cambrian “small shelly fauna”, archaeocyathans, early trilobites…) (Liñán & Palacios, 1987; Vidal et al., 1994; Liñán et al., 2004; Fernández-Remolar et al., 2005; Jensen et al., 2007; Zhuravlev et al., 2012; Jensen & Palacios, 2016). In the Central Iberian Zone (CIZ), the Ediacaran—Cambrian transition is recorded in several rock units which have received different names depending on the region and author (Nozal Martín et al., 1988; Vidal et al., 1994; Pieren Pidal, 2000). The present work deals with the Alcudian unit (Alcudiense, as described by Crespo & Tamain, 1971) in its typical region, the Alcudia Anticline of the valle de Alcudia (Ciudad Real, Sierra Morena), at the southern margin of the CIZ (Fig. 1; after Pieren Pidal, 2009).
|
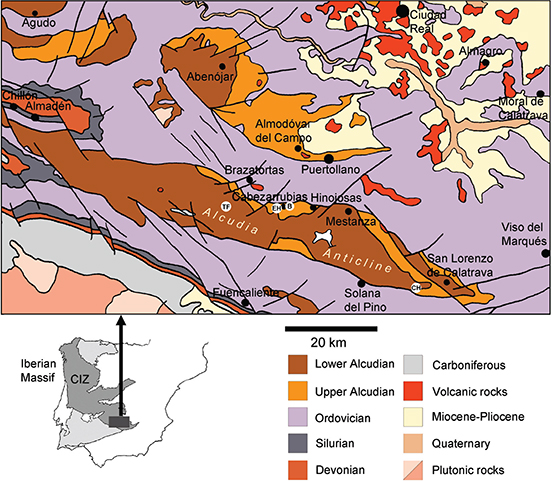 Fig. 1.—Geological map of the region of the Alcudia Anticline. CIZ = Central Iberian Zone. White circles indicate fossil sites described in the results: B = Arroyo del Barranco; TF = Tiesa Ferrer; EH = El Hontanar; CH = El Chorrillo. Main map redrawn after Pieren Pidal (2000, 2009). The trace fossils of Fig. 3 come from two localities within the circle EH (see the corresponding figure caption). Fig. 1.—Geological map of the region of the Alcudia Anticline. CIZ = Central Iberian Zone. White circles indicate fossil sites described in the results: B = Arroyo del Barranco; TF = Tiesa Ferrer; EH = El Hontanar; CH = El Chorrillo. Main map redrawn after Pieren Pidal (2000, 2009). The trace fossils of Fig. 3 come from two localities within the circle EH (see the corresponding figure caption).
|
|
The axis of the valle de Alcudia is the Lower Alcudian subunit, a thick (4,000–6,000 m), generally subvertical, usually flysch-like alternation of greywackes and siltstones representing turbidites and gravitational breccias of probable late Ediacaran age, being younger than about 580–560 million years (Ma) according to radiometric geochronology using detrital zircons (Talavera et al., 2015). This subunit is considered to have been deposited on a continental slope (Pieren Pidal, 2000; 2009). Most of the Lower Alcudian is so fractured and folded that conventional stratigraphic study seems impossible. Its rocks are weakly metamorphosed, corresponding to the low and occasionally high anchizone (Pieren Pidal, 2000; 2009). This subunit is overlain by the Upper Alcudian, in angular unconformity (< 550–540 Ma according to Talavera et al., 2015).
Upper Alcudian beds tend to be much less distorted, suffered less metamorphism, and contain a wide range of lithologies, with frequent conglomerates. In the Alcudia Anticline, this subunit was divided by Pieren Pidal & García-Hidalgo (1999) into five formations which are interpreted to record sedimentation in a shallow siliciclastic platform. From bottom to top, these formations (Fm.) are the Tamujar, Hinojosas, Cabezarrubias (Pizarras de Cabezarrubias), San Lorenzo (Conglomerados de San Lorenzo), and Upper Formation (Pizarras Superiores) (Fig. 2). The nature of their contacts is still quite open to debate. Carbonate lenses are present in the Hinojosas Fm. (Pieren Pidal, 2000, 2009).
|
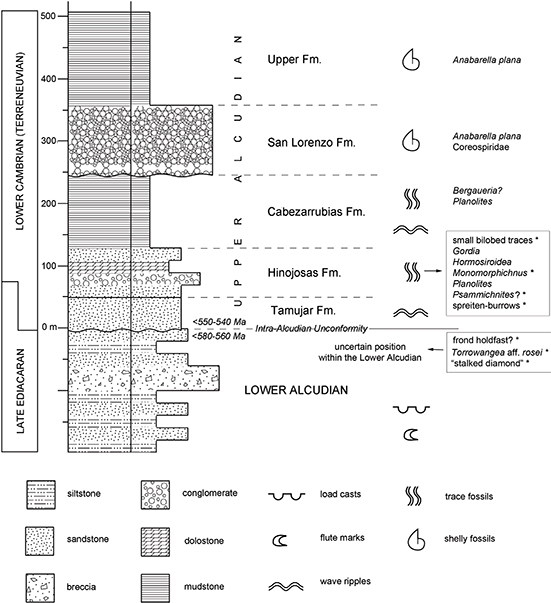 Fig. 2.—Simplified stratigraphic log of the Alcudian unit in the valle de Alcudia, showing only main rock types in each formation - bed sequence and thickness within each formation is arbitrary. Thickness of Upper Alcudian formations is approximately the same of the type section of each one, but the thickness of the Lower Alcudian is about 4,000-6,000 m. The asterisk indicates material described in the present work. Based on information from Pieren Pidal (2000; 2009). Fig. 2.—Simplified stratigraphic log of the Alcudian unit in the valle de Alcudia, showing only main rock types in each formation - bed sequence and thickness within each formation is arbitrary. Thickness of Upper Alcudian formations is approximately the same of the type section of each one, but the thickness of the Lower Alcudian is about 4,000-6,000 m. The asterisk indicates material described in the present work. Based on information from Pieren Pidal (2000; 2009).
|
|
Up to date, the fossil record of the Lower Alcudian is restricted to a single report of sphaeromorph acritarchs in an unpublished work by Mitrofanov & Timofeev (1979, cited from Pieren Pidal, 2009). The Upper Alcudian starts with hints of bioturbation in the Tamujar Fm. (Pieren Pidal, 2000). The Hinojosas Fm. has yielded the richest fossil assemblage of the Alcudia Anticline (García-Hidalgo, 1993; Pieren Pidal, 2000), consisting on varied trace fossils. In the northern flank (Cabezarrubias-Hinojosas zone, Fig. 1), the Hinojosas Fm. have simple unbranched burrows (Planolites), arthropod scratches (Monomorphichnus lineatus), and possible treptichnids assigned to Hormosiroidea canadensis. The presence of Monomorphichnus indicates a Cambrian age (Liñán et al., 1984; Gámez-Vintaned & Liñán, 1996, 2007). In the southern flank of the anticline (Macizo de Valdoro area), this formation contains simple traces with loops (Gordia molassica), and meandering bilobed traces initially identified as Taphrhelminthopsis but reinterpreted by Jensen & Palacios (2016) as possible Psammichnites circularis. However, according to the diagnosis of Taphrhelminthopsis, this ichnogenus refers to traces that are usually big (1–3 cm wide) and very long, whereas the bilobed trails of the Hinojosas Fm. are small (~4–5 mm wide) and not very long (see photograph 4 in Pieren Pidal, 2009). Thus the ichnogenus Archaeonassa (1–7 mm wide) seems more adequate for these trails (see the diagnoses of both ichnogenera in Häntzschel, 1975). An example of Archaeonassa from the upper Ediacaran of the White Sea region (figure 3 in Jensen, 2003) in fact looks very similar to these bilobed traces in size and path.
|
 Fig. 3.—El Hontanar trace fossil site. A. Exposure of mudstone beds; one of the upper bed tops contains the fossils illustrated from 3B to 3E. B. Oblique section of a spreiten-burrow showing some crescent-shaped units of a possible teichichnid. C. Monomorphichnus lineatus consisting on four scratch marks preserved as grooves in the bed top. D. Small meandering bilobed trail similar to Archaeonassa. E. Intense bioturbation showing trails (top) and burrows (centre). F. Bioturbation with many burrows of uncertain identification, sometimes with helicoidal-like distortion. Photograph F comes from a mudstone bed top about 75 meters north of the main outcrop, in the slope of the western bank of a stream that crosses the Hinojosas Fm. Field photographs. Scale bars = 1 cm. Fig. 3.—El Hontanar trace fossil site. A. Exposure of mudstone beds; one of the upper bed tops contains the fossils illustrated from 3B to 3E. B. Oblique section of a spreiten-burrow showing some crescent-shaped units of a possible teichichnid. C. Monomorphichnus lineatus consisting on four scratch marks preserved as grooves in the bed top. D. Small meandering bilobed trail similar to Archaeonassa. E. Intense bioturbation showing trails (top) and burrows (centre). F. Bioturbation with many burrows of uncertain identification, sometimes with helicoidal-like distortion. Photograph F comes from a mudstone bed top about 75 meters north of the main outcrop, in the slope of the western bank of a stream that crosses the Hinojosas Fm. Field photographs. Scale bars = 1 cm.
|
|
Above the Hinojosas Fm., possible cnidarian burrows (reported as Bergaueria aff. langi) and Planolites have been found in the Cabezarrubias Fm. The San Lorenzo Fm. contains the early mollusks Anabarella plana and undescribed members of the family Coreospiridae (previously Latouchellidae). Phosphatized remains of the Tommotian species Anabarella plana are extremely abundant in some massive mudstone beds of the Upper Fm. (Vidal et al., 1999; Pieren Pidal, 2009).
Material and methodsTOP
The specimens have been studied in outcrop when they appear in hard rocks and extensive beds that make them uncollectable without high risk of damage. They were photographed using a Canon EOS 550D and Canon EOS 7D DSLR cameras with a zoom lens Canon 18–55mm f3.5–5.6 ISII, and macro lenses Canon 60mm f2.8 and Canon 100mm f2.8. Collected specimens are deposited in the Museo de Paleontología de Castilla-La Mancha (Cuenca, Spain) under accession numbers MPCM-VA-0001 to MPCM-VA-0005.
ResultsTOP
Cambrian trace fossilsTOP
New Cambrian traces for the Alcudia Anticline were found in two localities. The first one is El Hontanar (Fig. 1), southwest of Cabezarrubias del Puerto (main surface at coordinates N 38º 36’ 44’’ W 4º 11’ 40’’), in the type section of the Hinojosas Fm. (Camino de Cotofía, Pieren Pidal, 2000, 2009). Most of the trace fossils appear in a bed top of a sequence of subhorizontal mudstone beds (Fig. 3A). Bioturbation was so intense (e.g. Fig. 3E, F) that the recognition of individual traces is difficult. However, three types of traces can be distinguished:
| I) |
Small bilobed trails: abundant, 2–3 mm wide, with a central furrow and two lateral ridges, sometimes meandering (as in Fig. 3D). Their small width and limited horizontal development suggest the ichnogenus Archaeonassa instead of the wider (1–3 cm) and mostly very long Taphrhelminthopsis (Häntzschel, 1975). |
| II) |
Series of subparallel grooves preserved in the bed top (Fig. 3C), interpreted as scratches of possible arthropods and assigned to Monomorphichnus lineatus (Crimes et al., 1977). There are several specimens in the main surface; the best preserved one is shown in Fig. 3C. |
| III) |
One vertical section of what appears to be an inclined burrow with spreite (Fig. 3B). It is about 4 cm long and contains crescent-shaped units of decreasing size and about 1 cm of maximum width. The trace is similar to Teichichnus, the simplest spreiten-burrow, but the poor preservation recommends no assignation further than possible teichichnid. |
The second trace fossil locality is the Arroyo del Barranco outcrop (Fig. 4), southeast of Cabezarrubias (N 38º 36’ 46’’ W 4º 10’ 40’’), in a section with carbonate lenses, again in the Hinojosas Fm. according to Pieren Pidal (2000). Next to the stream, a mudstone boulder was found with a surface plenty of burrows about 8 mm wide and sometimes more than 20 cm long. They are preserved as unilobed and unbranching positive reliefs whose infill shows very fine transverse striation in the best preserved stretches (Fig. 4B, arrow). The traces are predominantly horizontal and gently curved, and they often cross each other.
|
 Fig. 4.—Boulder with possible ventral surfaces of Psammichnites from the Arroyo del Barranco locality (MPCM-VA-0001). A. Field photograph of the burrow system. Lens cap diameter = 58 mm. B. Close-up of some burrows showing very fine transversal striation (arrow). Scale bar = 1 cm. Fig. 4.—Boulder with possible ventral surfaces of Psammichnites from the Arroyo del Barranco locality (MPCM-VA-0001). A. Field photograph of the burrow system. Lens cap diameter = 58 mm. B. Close-up of some burrows showing very fine transversal striation (arrow). Scale bar = 1 cm.
|
|
All these characteristics suggest the burrows represent the lowermost surface of Psammichnites (Seilacher, 2007). However, the ventral morphology of this ichnogenus can be highly variable and so it was not considered to be diagnostic by Mángano et al. (2002). In the emended diagnosis of Psammichnites, these authors referred to the bilobed upper surface only. Thus the traces of Fig. 4 do not meet the complete diagnosis of the ichnogenus, and so they are better described as very possible Psammichnites. The burrows resemble a smaller version of some specimens reported as Scolicia (but they are probably the ventral expression of Psammichnites) from the Lower Cambrian of the Azorejo Fm. in the Toledo Mountains (Moreno et al., 1976, their figures 3d and e).
Ediacaran structuresTOP
In Lower Alcudian beds, a number of structures were discovered which resemble Ediacara-type soft-bodied fossils, but their biogeneicity is problematic. A complex discoidal structure (Fig. 5B) was found in the sole of a coarse-grained turbidite immediately below the intra-Alcudian unconformity of El Chorrillo (Solanilla del Tamaral, Fig. 1) described by Palero (1993) and dated by Talavera et al. (2015), who found it to be younger than 580±7 Ma using detrital zircon radiometric geochronology. The disc is very similar to the holdfast of an Ediacaran frond such as Charniodiscus, but ultimate proofs of biogeneicity are lacking. The surface around it shows probable deep oscillation ripples but not a single similar disc was found in the outcrop.
|
 Fig. 5.—Late Ediacaran structures from the Alcudia Anticline. A. Turbidite outcrops crossed by the Tablillas river in the Tiesa Ferrer area. B. Discoidal hyporelief that could be a frond holdfast. C. Torrowangea aff. rosei (arrow) from the top of a thin silty turbidite coated with iron oxides (MPCM-VA-0002). D. Detail of Torrowangea aff. rosei from the previous image, showing small-scale meandering (lower arrow), and irregular constrictions originating pod-like segments (upper arrow and slightly above the lower arrow). E. Rimless concavity in a bed top, interpreted as a Longmyndian Beltanelliformis-like pseudofossil (MPCM-VA-0003). F. “Peanut blister” in a bed top (MPCM-VA-0004). Field photographs. Scale bars = 1 cm. Fig. 5.—Late Ediacaran structures from the Alcudia Anticline. A. Turbidite outcrops crossed by the Tablillas river in the Tiesa Ferrer area. B. Discoidal hyporelief that could be a frond holdfast. C. Torrowangea aff. rosei (arrow) from the top of a thin silty turbidite coated with iron oxides (MPCM-VA-0002). D. Detail of Torrowangea aff. rosei from the previous image, showing small-scale meandering (lower arrow), and irregular constrictions originating pod-like segments (upper arrow and slightly above the lower arrow). E. Rimless concavity in a bed top, interpreted as a Longmyndian Beltanelliformis-like pseudofossil (MPCM-VA-0003). F. “Peanut blister” in a bed top (MPCM-VA-0004). Field photographs. Scale bars = 1 cm.
|
|
The other structures come from the Tiesa Ferrer area (Almodóvar del Campo, Fig. 1), one of the least distorted exposures of the Lower Alcudian in the valle de Alcudia. These subvertical beds (Fig. 5A) contain silty Tc-d turbidites, 1–2 cm thick (Pieren Pidal, 2000), whose bedding planes are coated with iron oxides and show a texture similar to the Textured Organic Surfaces (TOS) typical of fossil-bearing beds in many Ediacaran sites (Gehling & Droser, 2009). This kind of surface and bed provided the object of Fig. 6, which was found in a debris pile immediately below a series of these turbidites (thus its way-up is not clear). It is the convex cast of a diamond-shaped sheet showing a diagonal groove apparently connected to a tubular structure. Some rings can be seen in this “stalk”. There are rib-like ridges extending from the diagonal axis outwards. The sheet display hints of an internal complex structure (serial branching?). The distinctive feature of the upper corner of the “diamond” suggests the sheet was folded backwards after it pressed the sediment surface. Overall, this “stalked diamond” appears to be the cast of a soft-bodied organism, but it would be highly unusual and, after extensive search, no similar objects were found in the outcrop.
|
 Fig. 6.—A possible soft-bodied fossil from the late Ediacaran of the Alcudia Anticline (MPCM-VA-0005). Upper arrow, imprint of a corner of the main sheet that was apparently folded backwards before lithification. Middle arrow, hints of internal structure. Lower arrow, tubular “stalk” with rings. Field photograph. Scale bar = 1 cm. Fig. 6.—A possible soft-bodied fossil from the late Ediacaran of the Alcudia Anticline (MPCM-VA-0005). Upper arrow, imprint of a corner of the main sheet that was apparently folded backwards before lithification. Middle arrow, hints of internal structure. Lower arrow, tubular “stalk” with rings. Field photograph. Scale bar = 1 cm.
|
|
In some instances, this type of thin turbidites contains filamentous structures in the TOS-like top surfaces (Fig. 5C-D, some other specimens examined in the field). They are tiny strings about 0.5–1 mm wide and up to 3 cm long, sometimes with small meanders (Fig. 5D, lower arrow) and irregular constrictions that eventually originate pod-like segments (see examples in the upper arrow of Fig. 5D and slightly above the lower arrow). These characteristics fit the diagnosis of the ichnogenus Torrowangea (Webby, 1970). The filaments are assigned to Torrowangea aff. rosei, a form described by Liñán & Palacios (1987) from late Ediacaran fine-grained beds of the Domo Extremeño Group in the nearby Cáceres province. Other occurrences of Torrowangea, also from the Domo Extremeño Group, come from the Estenilla and Cíjara Formations in the Río Huso section (Gámez-Vintaned, 1996; Jensen & Palacios, 2016). The interpretation of Torrowangea aff. rosei is problematic since it may be a simple trace fossil, a microbial filament, or the remains of a tubular organism (Jensen & Palacios, 2016). The presence of small-scale meandering in the present material could support a trace fossil interpretation, but additional specimens would be necessary to sustain this conclusion.
A significant microbial coating in these Ediacaran pre-turbidite bottoms is suggested by the finding of a number of ellipsoidal, rimless, concave epireliefs (Fig. 5E) very similar to some pseudofossils of the Longmyndian Group (England). Menon et al. (2015) demonstrated these Beltanelliformis-like concavities to be pseudofossils originated by the small-scale sinking of a sand cone after an upwards fluid injection obstructed by a microbial mat. Other structures suggesting substantial microbial development in the same kind of bed tops consist on “bubbles” (Fig. 5F) resembling the “peanut blisters” that Seilacher (2007) interpreted as gas escape structures in ancient microbial mats.
DiscussionTOP
The sedimentary environment inferred for the Cambrian trace fossils here described is the shallow platform where the Hinojosas Fm. was deposited (Pieren Pidal, 2000, 2009). The carbonates and abundant bioturbation point to low sedimentation rates. Age is constrained by the presence of Anabarella plana in overlying beds (Fig. 2). The age of this mollusk species ranges from about 535 to 531 Ma according to Gubanov & Peel (2003), which implies a Fortunian stage (Cambrian stage 1) for the ichnofossil locality, if the base of the stage 2 is at about 529 Ma as indicates the GSSP Table of the International Commision on Stratigraphy (consulted in June 5th 2017). In the regional chronology, this corresponds to the Lower Corduban substage (see its definition in Liñán et al., 2006).
The possible taxonomic affinities of the organisms which created the traces of El Hontanar and Arroyo del Barranco are varied. Monomorphichnus lineatus corresponds to arthropods or arthropod-like organisms (Crimes et al., 1977). As a possible teichichnid, the spreiten-burrow could have been excavated by annelid-grade vermiform creatures, or arthropods (Seilacher, 2007). In the original description of Archaeonassa, Fenton & Fenton (1937) showed that it closely resembles modern trails of the gastropod Littorina, and thus early mollusks (may be Anabarella plana or a similar species), or the mollusk-like halkieriids, could have produced the small bilobed trails of Fig. 3D. Seilacher & Gámez-Vintaned (1996) argued that the Psammichnites organism was an infaunal, soft-bodied, vermiform animal which bulldozed the sediment and had a “snorkel” appendage that protruded and collected food on the seafloor (Seilacher, 2007). The taxonomic affinity of such an organism is open to debate and may be close to mollusks or proboscis-bearing “worms”.
The Cambrian traces here described could belong to the lowermost Cambrian ichnozone (that is, the Treptichnus pedum + Monomorphichnus lineatus Zone in Spain, and the global and probably equivalent Treptichnus pedum Zone, see Narbonne et al., 1987; Gámez-Vintaned & Liñán, 2007). However, the time spans of these traces extend into younger zones, and so they could as well represent the second Cambrian ichnozone (the global Rusophycus avalonensis Zone and the very similar Spanish Rusophycus avalonensis + Rusophycus bonnarensis Zone; Narbonne et al., 1987; Gámez-Vintaned and Liñán, 2007). This is the ichnozone information provided by the Cambrian traces:
| • |
Monomorphichnus lineatus first appear in the lowermost Cambrian ichnozone (Liñán et al., 1984; Narbonne et al., 1987; Gámez-Vintaned & Liñán, 2007). |
| • |
The FAD of spreiten-burrows in Newfoundland and Spain (Teichichnus in both cases) is in the second Cambrian ichnozone (Narbonne et al.,1987; Gámez-Vintaned & Liñán, 2007). However, burrows with spreite of upper Ediacaran age have been found in the Nama Group (MacDonald et al., 2014). Thus the presence of a spreiten-burrow is not diagnostic of any Cambrian ichnozone. |
| • |
Similarly, although Archaeonassa was described in lower Cambrian rocks (Fenton and Fenton, 1937), it first appeared in the upper Ediacaran (Jensen, 2003). |
| • |
Psammichnites was reported by Gámez-Vintaned (1996) to occur a few meters above the possible Ediacaran—Cambrian boundary of the Río Huso section (CIZ), very close to the first apparition of Treptichnus pedum and Monomorphichnus lineatus and so in the lowermost Cambrian ichnozone. |
Regarding the Ediacaran structures here described, possible fossils comparable to the disc and the “stalked diamond” are not known for the late Ediacaran of Spain (Pieren Pidal, 2000; Jensen et al., 2007; however, see a possible occurrence of Nimbia in Liñán et al., 2009). These two structures remain as isolated findings in spite of extensive search, but rarity is not necessarily an argument against biogeneicity. The reason is that well exposed bedding planes are extremely rare in the Lower Alcudian, and deep marine complex organisms probably were scarce in the latest Ediacaran after the extinction of most rangeomorphs (Xiao & Laflamme, 2009), and so very few fossil findings would not be surprising in this unit. Biogeneicity seems more likely for the “stalked diamond” due to the very unusual arrangement of a ringed tubular structure with a quite symmetric sheet that displays ribs and hints of taphonomic folding and internal structure. This arrangement of different objects is very difficult to explain from an abiotic point of view. It may be a (degraded?) organism of uncertain taxonomic affinity, or an abiotic structure of unknown origin.
The occurrence of Torrowangea aff. rosei may represent the first finding of a trace fossil in the Lower Alcudian of the valle de Alcudia, if this kind of filamentous reliefs represented traces, as the possible meandering suggests. However, the options of microbial strings and tubular organisms cannot be definitely ruled out with this material. If they were trace fossils of bilateral organisms, they would imply that at least some parts of the Lower Alcudian are younger than about 555 Ma, since this is the age of the oldest unambiguous bilateral traces (Jensen, 2003; Fedonkin et al., 2007). This age would be in agreement with the radiometric dating of this subunit (< 580–560 Ma according to Talavera et al., 2015).
A trace fossil origin for these filaments is compatible with their occurrence as positive epireliefs, since Torrowangea could represent a string of faecal pellets (Liñán & Palacios, 1987). The deep marine environment is also coherent with the trace fossil hypothesis, because similarly simple traces have been described from turbidite outcrops that document the Ediacaran—Cambrian transition, for example in the Puncoviscana Formation (Buatois et al., 2014). More material is required to reach any solid conclusion about the origin of all these structures.
ACKNOWLEDGEMENTSTOP
To Jim Gehling for his encouragement, advice, and attention. To José Luis García-Hidalgo, Latha Menon, and especially Alex Liu, for their useful comments about some of the material here published. To Eladio Liñán and Sören Jensen, whose insightful reviews and advice served to improve the quality of this work. The specimens have been collected with permission from the heritage section of the Junta de Comunidades de Castilla-La Mancha (Exp. 170814-P1), under the project “Prospección de fósiles en rocas del Precámbrico y Cámbrico de Ciudad Real”.
ReferencesTOP
| ○ |
Buatois, L.A.; Narbonne, G.M.; Mángano, G.; Carmona, N.B. & Myrow, P. (2014). Ediacaran matground ecology persisted into the earliest Cambrian. Nature Communications, 3544. https://doi.org/10.1038/ncomms4544 |
| ○ |
Crespo, V.L. & Tamain, G. (1971). Mise en évidence de niveaux carbonatés interstratifiés dans l’Alcudien de la Sierra Morena centrale (Espagne). Comptes Rendus de l’Academie des Sciences. Série D. Sciences Naturelles, 272: 688–690. |
| ○ |
Crimes, T.P.; Legg, I.; Marcos, A. & Arboleya, M. (1977). Late Precambrian-low Lower Cambrian trace fossils from Spain. In: Trace Fossils 2 (Crimes, T.P. & Harper, J.C., Eds.), Geological Journal Special Issues 9, 91–138. |
| ○ |
Crimes, T.P. (1992). The record of trace fossils across the Proterozoic—Cambrian boundary. In: Origin and early evolution of the Metazoa (Lipps, J.H. & Signor, P.W., Eds.), Springer, New York, 172–202. https://doi.org/10.1007/978-1-4899-2427-8_6 |
| ○ |
Fedonkin, M.A.; Gehling, J.G.; Grey, K.; Narbonne, G.M. & Vickers-Rich, P. (2007). The rise of animals – Evolution and diversification of the kingdom Animalia. The Johns Hopkins University Press, Baltimore, 327 pp. |
| ○ |
Fenton, C.L. & Fenton, M.A. (1937). Archaeonassa: Cambrian snail trails and burrows. American Midland Naturalist, 18(3): 454–456. https://doi.org/10.2307/2420587 |
| ○ |
Fernández Remolar, D.C.; García-Hidalgo, J.F. & Moreno-Eiris, E. (2005). Interés del registro de los primeros organismos en el Arcaico y Proterozoico. Boletín de la Real Sociedad Española de Historia Natural, Sección Geológica, 100: 177–209. |
| ○ |
Gámez Vintaned, J.A. (1996). Field Trip Itineraries. II.1.b: The río Huso section. In: Field trip guide and abstracts, II Field Conference of the Cambrian Stage Subdivision Working Groups (Liñán, E.; Gámez Vintaned, J.A. & Gozalo, R., Eds.). Universidad de Zaragoza, Zaragoza, 28–31. |
| ○ |
Gámez Vintaned, J. A. & Liñán, E. (1996). Significant ichnological data during the Neoproterozoic-early Cambrian transition in Iberia. In: Field trip guide and abstracts, II Field Conference of the Cambrian Stage Subdivision Working Groups (Liñán, E.; Gámez Vintaned, J.A. & Gozalo, R., Eds.), Universidad de Zaragoza, Zaragoza, 101–102. |
| ○ |
Gámez Vintaned, J.A. & Liñán, E. (2007). The Precambrian—Cambrian boundary in Spain: ichnofossil palaeobiology and zonation. In: The Rise and Fall of the Vendian (Ediacaran) Biota. Origin of the Modern Biosphere (Semikhatov, M.A., Ed.), Geos, Moscow, 54–57. |
| ○ |
García Hidalgo, J.F.(1993). Las pistas fósiles de los anticlinales de Alcudia y Abenójar (Zona Centroibérica). Edad de las series. Geogaceta, 14: 57–59. |
| ○ |
Gehling, J. G. & Droser, M. L. (2009). Textured organic surfaces associated with the Ediacara biota in South Australia. Earth-Science Reviews, 96(3): 196–206. https://doi.org/10.1016/j.earscirev.2009.03.002 |
| ○ |
Gubanov, A. P. & Peel, J. S. (2003). The Early Cambrian helcionelloid mollusc Anabarella Vostokova. Palaeontology, 46(5): 1073–1087. https://doi.org/10.1111/1475-4983.00334 |
| ○ |
Häntzschel, W. (1975). Treatise on invertebrate paleontology, Supplement 1, Part W, Miscellanea, Trace fossils and problematica. Second edition (Revised and enlarged). The Geological Society of America and the University of Kansas, Boulder, Colorado. 269 pp. |
| ○ |
Jensen, S. (2003). The Proterozoic and earliest Cambrian trace fossil record; patterns, problems and perspectives. Integrative and Comparative Biology, 43(1): 219–228. https://doi.org/10.1093/icb/43.1.219 |
| ○ |
Jensen, S.; Palacios, T. & Mus, M.M. (2007). A brief review of the fossil record of the Ediacaran—Cambrian transition in the area of Montes de Toledo-Guadalupe, Spain. In: The rise and fall of the Ediacaran biota (Vickers-Rich, P. & Komarower, P., Eds.), Geological Society, London, Special Publications, 286: 223–235. https://doi.org/10.1144/sp286.16 |
| ○ |
Jensen, S. & Palacios, T. (2016). The Ediacaran—Cambrian trace fossil record in the Central Iberian Zone, Iberian Peninsula. Comunicaçoes Geologicas, 103: 83–92. |
| ○ |
Liñán, E.; Palacios, T. & Perejón, A. (1984). Precambrian-Cambrian boundary and correlation from southwestern and Central part of Spain. Geological Magazine, 121 (3): 221–228. |
| ○ |
Liñán, E. & Palacios, T. (1987). Asociaciones de pistas fósiles y microorganismos de pared orgánica del Proterozoico, en las facies esquisto grauváquicas del norte de Cáceres. Consecuencias regionales. Boletín de la Real Sociedad Española de Historia Natural (Geología), 84 (1–4): 211–232. |
| ○ |
Liñán, E.; Perejón, A.; Gozalo, R.; Moreno-Eiris, E. & Oliveira, J.T. (2004). The Cambrian system in Iberia. Instituto Geológico y Minero de España, Madrid. 63 pp. |
| ○ |
Liñán, E.; Gámez Vintaned, J.A.; Gozalo, R.; Dies, M.E. & Mayoral, E. (2006). Events and biostratigraphy in the Lower Cambrian of Iberia. Zeitschrift der deutschen Gesellschaft für Geowissenschaften, 157(4): 597–609. |
| ○ |
Liñán, E.; Gámez Vintaned, J.A. & Gozalo, R. (2009). Origen y diversificación de los animales pluricelulares. In: Paleontología de Invertebrados (Martínez Chacón, M.L. & Rivas, P., Eds.), Sociedad Española de Paleontología, 13–31. |
| ○ |
MacDonald, F.A.; Pruss, S.B. & Strauss, J.V. (2014). Trace fossils with spreiten from the late Ediacaran Nama Group, Namibia: complex feeding patterns five million years before the Precambrian–Cambrian boundary. Journal of Paleontology, 88(02): 299–308. https://doi.org/10.1666/13-042 |
| ○ |
Mángano, M.G.; Buatois, L.A. & Rindsberg, A.K. (2002). Carboniferous Psammichnites: systematic re-evaluation, taphonomy and autecology. Ichnos, 9: 1–22. https://doi.org/10.1080/10420940190034175 |
| ○ |
Menon, L.R.; McIlroy, D.; Liu, A.G. & Brasier, M. D. (2015). The dynamic influence of microbial mats on sediments: fluid escape and pseudofossil formation in the Ediacaran Longmyndian Supergroup, UK. Journal of the Geological Society, 173(1): 177–185. https://doi.org/10.1144/jgs2015-036 |
| ○ |
Moreno, F.; Vegas, R. & Marcos, A. (1976). Sobre la edad de las series ordovícicas y cámbricas relacionadas con la discordancia “Sárdica” en el anticlinal de Valdelacasa (Montes de Toledo, España). Breviora Geologica Asturica, 20(1): 8–15. |
| ○ |
Narbonne, G.M.; Myrow, P.M.; Landing, E. & Anderson, M.M. (1987). A candidate stratotype for the Precambrian-Cambrian boundary, Fortune Head, Burin Peninsula, southeastern Newfoundland. Canadian Journal of Earth Sciences, 24(7): 1277–1293. https://doi.org/10.1139/e87-124 |
| ○ |
Nozal Martín, F.; García Casquero, J.L. & Picart Boira, J. (1988). Discordancia Intraprecámbrica y series sedimentarias en el sector sur-oriental de los montes de Toledo. Boletín Geológico y Minero, 99(4): 473–489. |
| ○ |
Palero, F.J. (1993). Tectónica pre-hercínica de las series infraordovícicas del anticlinal de Alcudia y la discordancia intraprecámbrica en su parte oriental (Sector meridional de la Zona Centroibérica). Boletín Geológico y Minero, 104(3): 227–242. |
| ○ |
Pieren Pidal, A.P. (2000). Las sucesiones anteordovícicas de la región oriental de la provincia de Badajoz y área contigua de la de Ciudad Real. Universidad Complutense, Madrid. 620 pp. |
| ○ |
Pieren Pidal, A.P. (2009). Rasgos geológicos de la comarca de Puertollano y del valle de Alcudia (Ciudad Real, España). Memorias de la Real Sociedad Española de Historia Natural, 2ª época, 6: 95–132. |
| ○ |
Pieren Pidal, A.P. & García-Hidalgo, J.F. (1999). The Upper Alcudian from the Alcudia Anticline revisited (Ciudad Real, Central Spain). Journal of Conference Abstracts, 4(3):1018–1019. |
| ○ |
Seilacher, A. (2007). Trace fossil analysis. Springer, New York. 226 pp. |
| ○ |
Seilacher, A. & Gámez-Vintaned, J.A. (1996). Psammichnites gigas: una expression icnológica de la Explosión Cámbrica. XII Jornadas de Paleontología, Badajoz. 111–113. |
| ○ |
Talavera, C.; Martínez Poyatos, D. & González Lodeiro, F. (2015). SHRIMP U-Pb geochronological constraints on the timing of the intra-Alcudian (Cadomian) angular unconformity in the Central Iberian Zone (Iberian Massif, Spain). International Journal of Earth Sciences, 104: 1739-1757. https://doi.org/10.1007/s00531-015-1171-5 |
| ○ |
Vidal, G.; Palacios, T.; Moczydłowska, M. & Gubanov, A.P. (1999). Age constraints from small shelly fossils on the early Cambrian terminal Cadomian Phase in Iberia. GFF, 121(2): 137–143. https://doi.org/10.1080/11035899901212137 |
| ○ |
Vidal, G.; Palacios, T.; Gámez-Vintaned, J.A.; Díez Balda, M.A. & Grant, S.W.F.(1994). Neoproterozoic-early Cambrian geology and palaeontology of Iberia. Geological Magazine, 131(6): 729–765. https://doi.org/10.1017/S001675680001284X |
| ○ |
Webby, B.D. (1970). Late Precambrian trace fossils from New South Wales. Lethaia, 3(1): 79–109. https://doi.org/10.1111/j.1502-3931.1970.tb01265.x |
| ○ |
Xiao, S. & Laflamme, M. (2009). On the eve of animal radiation: phylogeny, ecology and evolution of the Ediacara biota. Trends in Ecology & Evolution, 24(1): 31–40. https://doi.org/10.1016/j.tree.2008.07.015 |
| ○ |
Zhuravlev, A.Yu.; Liñán, E.; Gámez Vintaned, J.A.; Debrenne, F. & Fedorov, A.B. (2012). New finds of skeletal fossils in the terminal Neoproterozoic of the Siberian Platform and Spain. Acta Palaeontologica Polonica, 57(1): 205–224. https://doi.org/10.4202/app.2010.0074 |