Los restos de Sus scrofa (Artiodactyla, Mammalia) del yacimiento Pleistoceno de Pinilla del Valle (Madrid, EspaÑa)
A study of Sus scrofa (Artiodactyla, Mammalia) from the Pleistocene fossil site of Pinilla del Valle (Madrid, Spain)
F. Alférez, A.M. Buitrago
1Departamento de Geodinámica, Estratigrafía y Paleontología, Facultad de Ciencias Geológicas, José Antonio Novais 12, 28040 Madrid. Email: alfdel3@yahoo.es, abuitrag05@gmail.com ORCID ID: https://orcid.org/0000-0001-5877-375X, http://orcid.org/0000-0001-9923-9668
| |
RESUMEN
Se ha llevado a cabo un estudio morfológico y biométrico de los restos fósiles de Suidos excavados en el yacimiento del Pleistoceno
superior de la Cueva del Camino, en el término de Pinilla del Valle (Madrid), comparándolos con los restos de otros yacimientos
y con especímenes actuales. El material extraído comprende 47 elementos pertenecientes a un mínimo de 6 individuos, destacando
la presencia de restos craneales, muy poco frecuentes en los yacimientos europeos de esta época, y ausentes en yacimientos
españoles. Se han comprobado en los restos los caracteres anatómicos típicos de Sus scrofa, confirmándose las condiciones paleoambientales obtenidas del conjunto de la fauna.
Palabras clave: Pleistoceno; MIS 5; yacimiento Cueva del Camino; primera fase de excavaciones; ungulados; Sus scrofa. |
| |
ABSTRACT
A morphologic and biometric study has been carried out on the fossil remains of Suidae derived from the first phase of excavation
(1980-1989) at the late Pleistocene paleontological site of Cueva del Camino near Pinilla del Valle, north of Madrid. The
material consists in 47 remains belonging to a minimum of 6 individuals. The results allow identifying the presence of the
wild boar Sus scrofa, whose anatomical characteristics are compared with material from various European Pleistocene sites as well as present-day
specimens. From the material obtained is this first phase of excavation stands out several cranial remains, because they are
very scarce in coeval European sites, and are not recorded so far in other Spanish sites. The record of Sus scrofa agrees with the palaeoecological and palaeoenvironmental inferences displayed by previous studies from the whole faunal assemblage
of the Cueva del Camino site.
Keywords: Pleistocene; MIS 5 record; Cueva del Camino site; first phase of excavation; Ungulates; Sus scrofa.
|
IntroducciónTOP
El yacimiento paleontológico de Pinilla del Valle se sitúa al norte de la provincia de Madrid, en la cabecera del río Lozoya
junto al embalse del mismo nombre, y en un promontorio calcáreo conocido como Calvero de las Sepulturas (actualmente redenominado
Calvero de la Higuera). Éste alberga uno de los yacimientos más conocidos del Cuaternario de esta Comunidad autónoma (Alférez et al., 1982, 1983; Maldonado, 1991; Arribas et al., 1998; Sesé & Soto, 2000, 2002; Corvea Porras et al., 2006; Pérez-González et al., 2010; Arsuaga et al. 2010, 2012; Martínez-Navarro et al., 2018), catalogado también entre los lugares de interés geológico españoles de relevancia internacional (Jordá Pardo, 2008; Carcavilla & Palacio, 2010) y que desde 2015 goza de reconocimiento y protección regional (Parque Arqueológico del Calvero de la Higuera: Baquedano et al., 2015).
El yacimiento principal corresponde a la denominada Cueva del Camino, una cavidad colapsada cuya edad de formación fue estimada
preliminarmente por Alférez et al. (1982), Carbó et al. (1990) y Alférez & Roldán (1992), como correspondiente a la parte alta del Pleistoceno medio (algo menos de 200 ka: interestadial Riss II-III). La datación
se basaba en la asociación faunística y en unos valores geocronométricos (obtenidos mediante resonancia paramagnética electrónica
–ESR– y series de uranio) de sendos espeleotemas estalagmíticos, que sin embargo no llegaron a ser publicados. Las modernas
dataciones con termoluminiscencia (Pérez-González et al., 2010) rebajaron notablemente la antigüedad del nivel fosilífero principal del yacimiento, estimándolo en 90,961 ± 7,761 ka BP.
Con ello, la asociación de mamíferos de la Cueva del Camino pasó de una adscripción provisional al Pleistoceno medio, a situarse
informalmente en el Pleistoceno superior. Este último calificativo se debe a que la Tabla Cronoestratigráfica Internacional
establece el inicio del Pleistoceno Superior a los 0,126 Ma (12,6 ka: Cohen et al., 2013, con actualización permanente en la página web de la International Commission on Stratigraphy/IUGS –www.ics.org–), una edad discordante con la asignada modernamente al yacimiento. Además y en términos de las escalas geocronológicas continentales,
la edad Eemiense apuntada por Alférez et al. (1982) para el yacimiento, se ha visto en cierto modo refrendada por datos nuevos derivados de los roedores y asociaciones específicas
de macromamíferos (Arsuaga et al., 2010, p. 431 y 436), toda vez que el Eemiense (o Riss-Würm) ha pasado a correlacionarse con la primera mitad del Pleistoceno superior
sensu lato (Estadio Isotópico Marino MIS 5) y a ser entendido, en su más amplio sentido, como el buen tiempo que precede a la Glaciación
Würm. De acuerdo con las modernas revisiones faunísticas y geocronométricas, a partir de Arsuaga et al. (2010) y Pérez-González et al. (2010), el yacimiento se considera coetáneo del último interglacial cálido, del periodo MIS 5 tardío, incluyendo el tránsito al
MIS 4 algo más frío (Blain et al., 2014).
El presente estudio se centra en la primera fase de excavaciones, desarrollada a lo largo de 10 años (1980-1989) bajo la dirección
del primero de los autores (FA), durante la que se extrajeron más de 3.000 restos de cinco clases de vertebrados, que fueron
identificados, siglados e inventariados. La asociación quedó tipificada por más de 70 especies pertenecientes a 16 órdenes
y 44 familias (Alférez & Molero, 1982; Alférez et al., 1982, 1985a, 1985b; Alférez, 1985, 1987; Alférez & Íñigo, 1990; Toñi & Molero, 1990; Buitrago, 1992; Maldonado, 1996; Bochenski, 2007). Entre los artiodáctilos no rumiantes, Alférez et al. (1982) identificaron tempranamente la presencia del jabalí Sus scrofa L. en la Cueva del Camino.
Esta abundancia de restos configura al yacimiento como uno de los más ricos del Pleistoceno español, correspondiendo además
a una época relativamente mal documentada en la Meseta española.
Entre los mamíferos hay una cantidad apreciable de suidos, teniendo en cuenta que, por regla general, los restos de este grupo
casi nunca son abundantes (o faltan) en los yacimientos del Pleistoceno medio y superior de Europa occidental.
Por otra parte, el material excavado tiene una gran importancia por los restos craneales encontrados, muy poco frecuentes
en los yacimientos europeos de esta época, y hasta ahora ausentes en los españoles.
Desde el punto de vista cronológico, los primeros fósiles ibéricos atribuidos al jabalí euroasiático corresponden a la identificación
de Sus sp. en el nivel TE9 de la Sima del Elefante (Atapuerca: Rodríguez et al., 2011), cuya edad se estima en 1,2 Ma (Carbonell et al., 2008), si bien hay autores que rebajan la misma a 0,9 Ma (Muttoni et al., 2013, 2018). A escala europea, los primeros jabalíes euroasiáticos bien caracterizados corresponden a la variedad Sus scrofa priscus Goldfuss (Guérin & Faure, 1997), según restos encontrados en el yacimiento alemán de Untermassfeld, próximo al millón de años de antigüedad (Kahlke, 2000).
Los registros previamente publicados de Sus scrofa en Pinilla se resumen en un M3 derecho ilustrado por Alférez et al. (1982, lám. 3, fig. 12, vista oclusal) y un fragmento de mandíbula derecha con la serie P3-M3 en vista lingual (Álvarez-Lao et al., 2013, fig. 3 m). Este último trabajo, centrado en los ungulados de la Cueva del Camino, destaca que la muestra de la especie,
obtenida en la segunda fase de las excavaciones desarrolladas en el yacimiento (2002-2009), es “demasiado pequeña para permitir
estudios comparativos” (tan sólo dos individuos, el 2% de la asociación de ungulados). No obstante, el material derivado de
la primera fase de las excavaciones es más amplio y menos fragmentario (Alférez & Buitrago, 1994), por lo que su estudio detallado, planteado en este trabajo, puede aportar datos de interés para el conocimiento de los
jabalíes del Pleistoceno europeo.
MaterialTOP
La mayoría de los restos de suidos excavados en el yacimiento son dentarios, incluyendo dos fragmentos craneales con la dentición
yugal casi completa en ambas maxilas; un fragmento maxilar derecho con P3–M3; un fragmento de maxilar juvenil con dentición de leche (DP2–M1), y otro pequeño fragmento con P2–P3 (Tabla 1). También hay tres mandíbulas incompletas y un fragmento mandibular, todos ellos con gran parte de la dentición (Tabla 2), y un cóndilo mandibular.
Tabla 1.—Dimensiones de los dientes deciduos y permanentes superiores.
| Sigla |
Elemento |
D |
DAP |
DT |
H |
DAPm |
DTm |
Hm |
| LPV-0226 |
DP2 drcho *. |
medio |
11,5 |
6,5 |
- |
- |
- |
- |
| LPV-0226 |
DP3 drcho *. |
medio |
10,4 |
10,4 |
- |
| LPV-0226 |
DP4 drcho *. |
medio |
15,0 |
13,3 |
- |
| LPV-2522 |
I1 drcho |
medio |
14,5 |
7,8 |
- |
- |
- |
- |
| LPV-1357 |
I2 drcho |
poco |
17,8 |
6,1 |
- |
| LPV-1686 |
P1 |
muy poco |
9,8 |
4,6 |
5,1 |
- |
- |
- |
| LPV-1399 |
P1 |
poco |
10,4 |
4,3 |
5,0 |
| LPV-1010 |
P2 drcho *. |
poco |
13,1 |
7,6 |
8,1 |
13,4
(n=4) |
7,5
(n=4) |
8,2
(n=4) |
| LPV-0798 |
P2 drcho *. |
poco |
14,3 |
7,6 |
8,0 |
| LPV-0798 |
P2 izqdo *. |
poco |
13,3 |
7,6 |
8,2 |
| LPV-1413 |
P2 izqdo *. |
poco |
13,2 |
7,3 |
8,5 |
| |
|
poco |
|
|
|
|
|
|
| LPV-1010 |
P3 drcho *. |
poco |
14,4 |
11,7 |
10,3 |
14,71
(n=7) |
12,01
(n=7) |
9,92
(n=7) |
| LPV-0798 |
P3 drcho *. |
poco |
15,0 |
12,5 |
10,0 |
| LPV-0798 |
P3 izqdo *. |
poco |
15,0 |
11,7 |
9,6 |
| LPV-1214 |
P3 drcho *. |
poco |
14,4 |
11,7 |
9,9 |
| LPV-1413 |
P3 izqdo *. |
poco |
14,1 |
11,3 |
9,8 |
| LPV-0321 |
P3 drcho |
poco |
15,2 |
12,7 |
9,9 |
| LPV-0430 |
P3 izqdo |
poco |
14,9 |
12,5 |
10,0 |
| LPV-1010 |
P4 drcho *. |
poco |
13,7 |
14,8 |
10,0 |
13,98
(n=5) |
15,66
(n=5) |
10,16
(n=5) |
| LPV-1010 |
P4 izqdo *. |
poco |
13,9 |
15,0 |
10,3 |
| LPV-0798 |
P4 drcho *. |
poco |
14,6 |
16,7 |
10,3 |
| LPV-0798 |
P4 izqdo *. |
poco |
13,9 |
15,0 |
10,2 |
| LPV-1214 |
P4 drcho *. |
poco |
13,2 |
15,5 |
10,0 |
| LPV-1010 |
M1 drcho *. |
medio |
19,1 |
16,6 |
- |
18,65
(n=7) |
16,33
(n=6) |
- |
| LPV-1010 |
M1 izqdo *. |
medio |
19,4 |
16,3 |
- |
| LPV-0798 |
M1 drcho *. |
medio |
17,6 |
- |
- |
| LPV-0798 |
M1 izqdo *. |
medio |
18,3 |
16,9 |
- |
| LPV-1214 |
M1 drcho *. |
medio |
18,4 |
16,6 |
- |
| LPV-1226 |
M1 drcho *. |
poco |
18,3 |
16,1 |
- |
| LPV-1692 |
M1 drcho |
sin gastar |
19,5 |
15,5 |
- |
| LPV-1010 |
M2 drcho *. |
poco |
25,1 |
21,4 |
12,3 |
25,4
(n=5) |
21,64
(n=5) |
12,18
(n=5) |
| LPV-1010 |
M2 izqdo *. |
poco |
25,6 |
21,4 |
12,7 |
| LPV-0798 |
M2 drcho *. |
poco |
25,7 |
22,2 |
11,6 |
| LPV-0798 |
M2 izqdo *. |
poco |
25,6 |
22,1 |
12,1 |
| LPV-1214 |
M2 drcho *. |
medio |
25,0 |
21,1 |
12,2 |
| LPV-1010 |
M3 drcho *. |
poco |
39,8 |
23,0 |
13,0 |
40,45
(n=6) |
23,71
(n=6) |
13,40
(n=5) |
| LPV-1010 |
M3 izqdo *. |
poco |
39,8 |
22,8 |
13,1 |
| LPV-0798 |
M3 drcho *. |
poco |
42,7 |
24,7 |
13,0 |
| LPV-0798 |
M3 izqdo *. |
poco |
41,2 |
24,5 |
13,7 |
| LPV-1214 |
M3 drcho *. |
poco |
39,6 |
23,1 |
14,2 |
| LPV-0320 |
M3 drcho |
medio |
39,6 |
24,2 |
- |
| Glosario de términos y abreviaturas (orden alfabético): ♀/♂ = Hembra/Macho; * = Elemento inserto en maxilar/mandíbula; A = Anchura; a/p = Anterior/posterior; art = Articular; C
= Canino; Cdi = C. Distal; Cla = C. Labial; Cli = C. Lingual; D = Desgaste; DAP = Diámetro antero-posterior; DI(1-2)/(1-2)= Incisivo deciduo (1-2)superior/(1-2)inferior; DP(1-4)/(1-4)= Premolar deciduo (1-4)superior/(1-4)inferior; DT = Diámetro transversal; H = Altura; I(1-2)/(1-2)= Incisivo (1-2)superior/(1-2)inferior; L = Longitud corona (labial); M(1-3)/(1-3)= Molar (1-3)superior/(1-3)inferior; m = Media; mx. = Máximo; P = Perímetro; P(1-4)/(1-4)= Premolar (1-4)superior/(1-4)inferior; p/m/d = Proximal/Medio/Distal; Re = Radio de curvatura externo; Ri = Radio de curvatura interno; S/I = Superior/Inferior;
t = total. |
Tabla 2.—Dimensiones de los dientes deciduos y permanentes inferiores. Abreviaturas en Tabla 1.
| Sigla |
Elemento |
D |
DAP |
DT (t) o (a/p) |
H |
L |
DAPm |
DTm (t) o (a/p) |
Hm |
Lm |
| LPV-1589 |
DI1 izqdo |
germen |
5,2 |
4,3 |
- |
17,5 |
- |
- |
- |
- |
| LPV-1383 |
I2 drcho |
medio |
12,1 |
8,6 |
- |
33,8 |
- |
- |
- |
- |
| LPV-2172 |
I2 izqdo |
medio |
10,9 |
8,4 |
- |
37,5 |
| LPV-1167 |
I2 drcho |
- |
11,7 |
8,8 |
- |
- |
| LPV-2691 |
I3 drcho |
medio |
7,7 |
5,6 |
- |
18,4 |
7,9
(n=4) |
5,95
(n=4) |
- |
18,6
(n=4) |
| LPV-0028 |
I3 drcho |
sin gastar |
7,7 |
5,9 |
- |
18,6 |
| LPV-0966 |
I3 izqdo |
medio |
8,0 |
5,9 |
- |
18,1 |
| LPV-1564 |
I3 izqdo |
sin gastar |
8,2 |
6,4 |
- |
19,3 |
| LPV-0315 |
P1 drcho * |
poco |
9,5 |
4,4 |
9,0 |
- |
9,60
(n=6) |
4,68
(n=6) |
8,50
(n=4) |
- |
| LPV-0315 |
P1 izqdo * |
poco |
9,2 |
4,5 |
- |
- |
| LPV-2512 |
P1 drcho |
- |
9,6 |
5,1 |
- |
- |
| LPV-2512 |
P1 izqdo * |
sin gastar |
9,6 |
5,0 |
7,8 |
- |
| LPV-2792 |
P1 izqdo * |
sin gastar |
9,5 |
4,8 |
8,4 |
- |
| LPV-0049 |
P1 izqdo * |
sin gastar |
10,2 |
4,3 |
8,8 |
- |
| LPV-0315 |
P2 drcho * |
poco |
11,1 |
5,8 |
9,0 |
- |
11,4
(n=4) |
6,02
(n=4) |
8,97
(n=4) |
- |
| LPV-2512 |
P2 drcho * |
poco |
11,3 |
6,1 |
8,9 |
- |
| LPV-2512 |
P2 izqdo * |
poco |
11,6 |
6,5 |
9,1 |
- |
| LPV-1009 |
P2 izqdo * |
poco |
11,6 |
5,7 |
8,9 |
- |
| LPV-0315 |
P3 drcho * |
poco |
14,4 |
7,5 |
10,5 |
- |
14,42
(n=6) |
7,84
(n=6) |
10,27
(n=6) |
- |
| LPV-0315 |
P3 izqdo * |
poco |
13,5 |
7,4 |
9,0 |
- |
| LPV-2512 |
P3 drcho * |
poco |
14,0 |
8,1 |
9,0 |
- |
| LPV-2512 |
P3 izqdo * |
poco |
14,6 |
8,1 |
11,1 |
- |
| LPV-1009 |
P3 drcho * |
poco |
14,8 |
- |
9,7 |
- |
| LPV-1178 |
P3 izqdo * |
sin gastar |
15,2 |
8,1 |
12,3 |
- |
| LPV-0315 |
P4 drcho * |
poco |
17,4 |
10,6 |
11,3 |
- |
16,31
(n=7) |
10,91
(n=7) |
11,83
(n=3) |
- |
| LPV-0315 |
P4 izqdo * |
poco |
17,4 |
10,9 |
11,7 |
- |
| LPV-2512 |
P4 drcho * |
poco |
15,5 |
10,7 |
- |
- |
| LPV-2512 |
P4 izqdo * |
poco |
15,5 |
10,9 |
- |
- |
| LPV-1009 |
P4 drcho * |
poco |
16,2 |
11,2 |
- |
- |
| LPV-1009 |
P4 izqdo * |
poco |
15,7 |
11,0 |
- |
- |
| LPV-1213 |
P4 izqdo * |
poco |
16,5 |
11,1 |
12,5 |
- |
| LPV-0315 |
M1 drcho * |
medio |
17,4 |
13,0 |
- |
- |
17,31
(n=7) |
13,24
(n=7) |
- |
- |
| LPV-0315 |
M1 izqdo * |
medio |
17,2 |
12,9 |
- |
- |
| LPV-2512 |
M1 drcho * |
medio |
17,2 |
13,9 |
- |
- |
| LPV-2512 |
M1 izqdo * |
medio |
17,3 |
13,7 |
- |
- |
| LPV-1009 |
M1 drcho * |
bastante |
17,2 |
13,2 |
- |
- |
| LPV-1009 |
M1 izqdo * |
bastante |
17,2 |
13,0 |
- |
- |
| LPV-1213 |
M1 izqdo * |
medio |
17,7 |
13,0 |
- |
- |
| LPV-0315 |
M2 drcho * |
poco |
25,8 |
16,6/17,0 |
- |
- |
24,78
(n=9) |
16,21/
17,45
(n=9) |
- |
- |
| LPV-0315 |
M2 izqdo * |
poco |
25,8 |
16,4/17,3 |
- |
- |
| LPV-2512 |
M2 drcho * |
medio |
24,3 |
16,3/18,0 |
- |
- |
| LPV-2512 |
M2 izqdo * |
medio |
24,7 |
15,7/18,0 |
- |
- |
| LPV-1009 |
M2 drcho * |
medio |
23,7 |
16,1/17,1 |
- |
- |
| LPV-1009 |
M2 izqdo * |
medio |
23,6 |
16,2/17,4 |
- |
- |
| LPV-1213 |
M2 izqdo * |
poco |
25,0 |
16,4/17,5 |
- |
- |
| LPV-1228 |
M2 izqdo |
sin gastar |
25,2 |
16,0/17,2 |
- |
- |
| LPV-1534 |
M2 izqdo * |
poco |
25,0 |
16,2/17,6 |
- |
- |
| LPV-0315 |
M3 drcho * |
poco |
40,7 |
19,0/18,7/14,4 |
15,7 |
- |
41,40 (n=7) |
19,27/
18,71/
14,92
(n=8/7/8) |
15,7
(n=3) |
- |
| LPV-0315 |
M3 izqdo * |
poco |
41,0 |
19,6/18,9/15,0 |
15,9 |
- |
| LPV-2512 |
M3 drcho * |
medio |
40,6 |
19,2/19,1/15,0 |
- |
- |
| LPV-2512 |
M3 izqdo * |
medio |
41,6 |
19,3/18,7/15,2 |
- |
- |
| LPV-1009 |
M3 drcho * |
poco |
43,6 |
18,6/18,0/15,0 |
- |
- |
| LPV-1009 |
M3 izqdo * |
poco |
- |
18,8/-/16,0 |
- |
- |
| LPV-1213 |
M3 izqdo * |
poco |
40,8 |
19,1/18,6/14,4 |
15,4 |
- |
| LPV-2468 |
M3 izqdo |
sin gastar |
41,5 |
20,6/19,0/14,4 |
15,8 |
- |
Tabla 3.—Dimensiones de los caninos superiores e inferiores. Abreviaturas en Tabla 1.
| Sigla |
Elemento |
S/I |
♀/♂ |
D |
Cli |
Cla |
Cd |
DAP |
DT |
H |
L |
P |
Re |
Ri |
Clim |
Clam |
Cdm |
DAPm |
DTm |
| LPV-0524 |
C drcho |
S |
|
ápice roto |
- |
- |
- |
31,5 |
21,6 |
˃ 59,5 |
˃ 113 |
87 |
40 |
23 |
- |
- |
- |
- |
- |
| LPV-2257 |
C drcho |
S |
|
Sin gastar |
- |
- |
- |
27,7 |
17,7 |
48,3 |
100 |
76,5 |
35 |
27 |
| LPV-1009 |
C drcho * |
I |
♀ |
- |
16,0 |
12,3 |
- |
17,0 |
9,0 |
30,0 |
- |
- |
35,5 |
- |
- |
- |
- |
- |
- |
| LPV-1009 |
C izqdo * |
I |
♀ |
- |
15,2 |
- |
11,7 |
17,5 |
8,5 |
28,0 |
- |
- |
32,0 |
- |
| |
|
|
|
- |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| LPV-0315 |
C drcho * |
I |
♂ |
- |
20,1 |
15,0 |
15,2 |
22,4 |
16,0 |
55,2 |
- |
- |
44,7 |
- |
23,12
(n=5) |
17,04
(n=5) |
17,74
(n=5) |
24,54
(n=5) |
15,28
(n=5) |
| LPV-0315 |
C izqdo * |
I |
♂ |
- |
20,8 |
15,0 |
15,3 |
22,1 |
16,5 |
58,6 |
- |
- |
47,0 |
- |
| LPV-2512 |
C drcho * |
I |
♂ |
- |
25,1 |
19,0 |
20,0 |
26,2 |
15,9 |
45,0 |
- |
- |
52,0 |
- |
| LPV-2512 |
C izqdo * |
I |
♂ |
- |
25,0 |
19,2 |
20,5 |
27,0 |
16,5 |
44,0 |
- |
- |
44,0 |
- |
| LPV-2209 |
C izqdo * |
I |
♂ |
- |
24,6 |
18,0 |
17,7 |
25,0 |
17,5 |
107,0 |
- |
- |
73,0 |
59,0 |
Tabla 4.—Dentición presente y medidas de los segmentos dentarios maxilares (superiores) y mandibulares (inferiores). Abreviaturas
en Tabla 1.
| Maxilares (dientes superiores) |
| Sigla |
Elemento |
L DP2-DP4 |
L P2-P3 |
L P2-P4 |
L M1-M3 |
Lm DP2-DP4 |
Lm P2-P3 |
Lm P2-P4 |
Lm M1-M3 |
| |
| LPV-1010 |
P2–M3 drcho * |
- |
27,6 |
41,2 |
78,6 |
38,8
(n=1) |
28,5
(n=4) |
42,2
(n=3) |
80,3
(n=5) |
| LPV-1010 |
P4–M3 izqdo * |
- |
- |
- |
78,6 |
| LPV-0798 |
P2–M3 drcho * |
- |
29,2 |
43,6 |
81,1 |
| LPV-0798 |
P2–M3 izqdo * |
- |
28,5 |
41,9 |
82,1 |
| LPV-1214 |
P2–M3 drcho |
- |
- |
- |
81,0 |
| LPV-1226 |
DP2-M1 drcho |
38,8 |
- |
- |
- |
| LPV-1413 |
P2–P3 izqdo |
- |
28,5 |
- |
- |
| |
| Mandíbulas (dientes inferiores) |
| Sigla |
Elemento |
L P1-P4 |
L P2-P4 |
L P3-P4 |
L P2-M3 |
L M1-M3 |
Lm P1-P4 |
Lm P2-P4 |
Lm P3-P4 |
Lm P2-M3 |
Lm M1-M3 |
| |
| LPV-0315 |
C-M3 drcho * |
69,6 |
43,1 |
31,8 |
125,5 |
83,7 |
71,63
(n=4) |
42,0
(n=3) |
30,85
(n=6) |
124,0
(n=3) |
83,05
(n=7) |
| LPV-0315 |
C–P1; P3-M3 izqdo * |
71,9 |
- |
31,8 |
- |
83,3 |
| LPV-2512 |
I1–M3 drcho |
73,0 |
41,6 |
29,5 |
122,7 |
82,5 |
| LPV-2512 |
I1–M3 izqdo |
72,0 |
41,4 |
30,1 |
123,8 |
83,9 |
| LPV-1009 |
C;P3-M3 drcho |
- |
- |
31,0 |
- |
83,3 |
| LPV-1009 |
C;P2;P4-M3 izqdo |
- |
- |
30,9 |
- |
83,3 |
| LPV-1213 |
P4-M3 izqdo |
- |
- |
- |
- |
82,4 |
| |
| Medidas de los diastemas y distancias desde el canino en las mandíbulas inferiores |
| Sigla |
Elemento |
L dias. C-P1 |
L dias. P1-P2 |
L C–P2 |
L P2-Cmesial |
L P1-Cmesial |
A máx. sínfisis |
Lm C-P1 |
Lm P1-P2 |
Lm C–P2 |
Lm P2-Cmesial |
Lm P1-Cmesial |
Am sínfisis |
| |
| LPV-0315 |
drcho * |
8,0 |
21,0 |
36,6 |
56,6 |
27,8 |
57,7 |
9,7
(n=4) |
19,7
(n=5) |
36,1
(n=4) |
59,4
(n=5) |
28,7
(n=4) |
54,9
(n=3) |
| LPV-0315 |
izqdo * |
8,0 |
20,3 |
35,7 |
56,0 |
27,2 |
| LPV-2512 |
drcho |
- |
19,2 |
- |
62,0 |
31,3 |
51,2 |
| LPV-2512 |
izqdo |
13,3 |
18,4 |
36,0 |
63,0 |
- |
| LPV-1009 |
drcho |
- |
- |
- |
- |
- |
55,9 |
| LPV-1009 |
izqdo |
8,0 |
21,0 |
36,6 |
56,6 |
27,8 |
Se han encontrado además 25 dientes aislados: 1 I1; 1 I2; 2 CS; 2 P1; 2 P3; 1 M1; 1 M3; 1DI1; 3 I2; 4 I3; 1 Ci; 2 P1; 1 P3; 2 M2 y 1 M3.
Los dientes implantados en los fragmentos craneales y mandibulares suman 87, lo que junto a los 25 dientes sueltos hacen un
total de 112 piezas dentarias las que han sido utilizadas en este estudio.
Los restos postcraneales encontrados son escasos y corresponden a una costilla, un fragmento distal de tibia, un navicular
y un metatarsiano III.
Este conjunto de restos corresponde a un número mínimo de 6 individuos (presencia de seis molares terceros inferiores izquierdos),
a los que habría que sumar los 24 elementos no especificados de dentición y fragmentos postcraneales mencionados por Álvarez-Lao et al. (2013), quienes cifran en un mínimo de dos, los individuos suplementarios encontrados durante los postreros ocho años de excavaciones
en la Cueva del Camino.
Las dimensiones normalizadas de los elementos fósiles estudiados, y su comparativa con los materiales de otros yacimientos
y procedencias se recogen en las tablas 5 y 7.
Tabla 5.—Dimensiones comparadas de dientes y segmentos dentarios de Pinilla del Valle con otros yacimientos y material actual.
| Elemento |
Medida |
Pinilla |
Jaurens |
Pl. m. |
Pl. s. |
Fage |
Cova Negra |
Casar. |
Mariz. |
Sussen. |
Ekain |
Europaact. |
Burgos 1-2 act. |
Pardo |
| Maxilares (dientes superiores) |
| |
|
LPV-0524
drcho |
LPV-2257
drcho |
|
|
|
|
|
|
|
|
|
|
|
|
| C |
DAP |
31,5 |
27,7 |
30,0 |
- |
- |
32,8 |
- |
- |
- |
- |
- |
- |
- |
22,5♂ |
| DT |
21,6 |
17,7 |
20,0 |
- |
- |
21,6 |
- |
- |
- |
- |
- |
- |
- |
16,0♂ |
| L |
˃113,0 |
100,0 |
110,0 |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
95,0♂ |
| P |
87,0 |
76,5 |
85,0 |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
58,0♂ |
| A |
˃59,5 |
48,3 |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
45,0♂ |
| Re |
40,0 |
35,0 |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
35,0♂ |
| M1 |
DAP |
18,6 |
|
18,5 |
19,0 |
17,2 |
- |
18,2 |
- |
- |
- |
- |
- |
15,8(1) |
- |
| DT |
16,3 |
|
17,0 |
15,5 |
14,9 |
- |
- |
- |
- |
- |
- |
- |
14,1(1) |
- |
| M2 |
DAP |
25,4 |
|
24,0 |
25,1 |
24,6 |
- |
- |
23,4 |
25,3 |
- |
- |
- |
23,4(1) |
22,5 |
| DT |
21,6 |
|
22,0 |
21,3 |
19,2 |
- |
- |
17,9 |
- |
- |
- |
- |
16,9(1) |
17,4 |
| M3 |
DAP |
40,4 |
|
39,0 |
- |
- |
- |
- |
34,5 |
35,4 |
- |
- |
- |
31,8(1) |
34,7 |
| DT |
23,7 |
|
22,0 |
- |
- |
- |
- |
20,0 |
- |
- |
- |
- |
19,1(1) |
19,1 |
| M1-M3 |
L |
80,3 |
|
- |
- |
- |
|
- |
- |
- |
|
|
|
72,0(1) |
71,0 |
| |
| Mandíbula (dientes inferiores) |
| |
|
LPV-2209 |
- |
|
|
|
|
|
|
|
|
|
|
|
|
| |
| C |
DAP |
25,0 |
|
26,0 |
13,5 |
19,8 |
- |
- |
- |
- |
- |
- |
21,8 |
- |
- |
| DT |
17,5 |
|
28,0 |
8,8 |
12,3 |
- |
- |
- |
- |
- |
- |
12,9 |
- |
- |
| P1 |
DAP |
9,6 |
|
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
7,8(1)♀ |
- |
| DT |
4,7 |
|
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
4,2(1)♀ |
- |
| P2 |
DAP |
11,4 |
|
14,5 |
13,4 |
11,9 |
- |
- |
- |
- |
- |
- |
12,1 |
12,4(1)♀ |
- |
| DT |
6,0 |
|
6,5 |
5,4 |
6,0 |
- |
- |
- |
- |
- |
- |
5,4 |
5,4(1)♀ |
- |
| P3 |
DAP |
14,4 |
|
15,5 |
14,8 |
13,8 |
- |
- |
- |
- |
- |
- |
13,2 |
14,0(1)♀ |
- |
| DT |
7,8 |
|
8,5 |
7,0 |
7,4 |
- |
- |
- |
- |
- |
- |
7,4 |
7,0(1)♀ |
- |
| P4 |
DAP |
16,3 |
|
15,5 |
15,9 |
15,0 |
- |
- |
- |
- |
- |
- |
14,2 |
14,7(1)♀ |
- |
| DT |
10,9 |
|
11,0 |
8,9 |
9,9 |
- |
- |
- |
- |
- |
- |
9,4 |
8,5(1)♀ |
- |
| M1 |
DAP |
17,3 |
|
17,0 |
16,2 |
16,7 |
- |
- |
- |
- |
- |
17,5 |
15,9 |
15,5(1)♀ |
- |
| DT |
13,2 |
|
14,0 |
11,8 |
11,9 |
- |
- |
- |
- |
- |
11,0 |
12,0 |
11,3(1)♀ |
- |
| M2 |
DAP |
24,8 |
|
23,0 |
23,5 |
21,7 |
- |
- |
- |
- |
20,5 |
- |
21,1 |
21,5(1)♀ |
- |
| DT (a/p) |
16,2/
17,5 |
|
17,0/
c17,0 |
15,7/
16,0 |
14,4/
14,9 |
- |
- |
- |
- |
16,0/ 17,0 |
- |
14,5/ 15,3 |
13,5(1)/
15,0(1)♀ |
- |
| M3 |
DAP |
41,4 |
|
42,0 |
38,9 |
36,8 |
- |
- |
- |
- |
- |
38,5 |
36,6 |
38,2(1)♀ |
- |
| DT (a/m/p) |
19,3/
18,7/
14,9 |
|
21,0/
19,0/
16,5 |
18,5/
14,9/- |
17,2/
13,2/- |
- |
- |
- |
- |
20,0/-/- |
16,3/-/- |
17,2/
14,8/
15,1 |
15,6(1)/
16,4(1)/
13,5(1)♀ |
- |
| P1–P4 |
L |
71,6 |
|
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
59,5(2) |
- |
| P2–P4 |
L |
420 |
|
450 |
43 |
40 |
49,4 |
- |
- |
- |
- |
- |
39,1 |
40,5(2) |
- |
| P3–P4 |
L |
30,8 |
|
31,0 |
29,6 |
27,6 |
- |
- |
- |
- |
- |
- |
27,4 |
28,0(2) |
- |
| P2-M3 |
L |
124,0 |
|
125,0 |
117,5 |
111,0 |
- |
- |
- |
- |
- |
- |
111,1 |
115,1(2) |
- |
| M1-M3 |
L |
83,0 |
|
81,0 |
76,0 |
72,4 |
- |
- |
- |
- |
- |
- |
72,0 |
75,0(2) |
- |
| Yacimientos de procedencia y otro material de comparación: Jaurens (Faure & Guérin, 1983), Monsbach y Mauer (Pl. m., Pleistoceno medio, Faure & Guérin, 1983), Pleistoceno superior y Holoceno (Pl. s., Faure & Guérin, 1983), La Fage (Fage, Bouchud, 1972), Cova Negra (Pérez Ripoll, 1977), Los Casares (Casar., Altuna, 1973), Marizulo (Mariz., Altuna, 1967), Sussenborg (Sussen., Kalhke, 1965), Ekain (Altuna & Mariezkurrena, 1984); más ejemplares actuales de Europa (Europa act., Faure & Guérin, 1983), Burgos (1, Burgos 1 act., col. MNCN, Madrid; 2, Burgos 2 act., Faure & Guérin, 1983) y El Pardo (Pardo, col. UCM, Madrid). Abreviaturas en Tabla 1. |
Tabla 6.—Dimensiones de los elementos postcraneales del esqueleto (la orientación se ha hecho con respecto al eje de la extremidad).
Abreviaturas en Tabla 1.
| Sigla |
Elemento |
DAP (t/art) o (p/m/d) |
DT (t/art) o (p/m/d) |
H (t) o (t/a) |
L |
DAPm |
DTm (t) o (p/m/d) |
Hm |
| LPV-1743 |
Tibia drcha |
32,6/28,5 |
34,8/27,9 |
- |
- |
- |
- |
- |
| LPV-1507 |
Navicular izqdo |
32,7/- |
19,2/- |
21,5/15,2 |
- |
- |
- |
- |
| LPV-2739 |
Metatarsiano III drcho |
28,4/15,7/19,4 |
21,5/16,6/19,7 |
- |
107,8 |
- |
- |
- |
Tabla 7.—Dimensiones comparadas de los elementos postcraneales del esqueleto de Pinilla del Valle con otros yacimientos y
material actual.
| Medida |
Pinilla |
Jaurens |
|
Marizulo |
Burgos (1) |
Burgos (3) |
Lyon |
Basilea |
| Tibia |
| |
LPV-1743 drcho |
|
|
|
|
|
drcho |
|
|
| DAP (t/art) |
32,6/28,5 |
- |
|
- |
- |
|
28,6/25,7♂ |
- |
- |
| DT (t/art) |
34,8/27,9 |
- |
|
- |
- |
|
33,0/24,0♂ |
- |
- |
| Navicular |
| |
LPV-1507 izqdo |
395044 izqdo |
395045 drcho |
|
izqdo |
drcho |
drcho |
22 drcho |
10687 izqdo |
| DAP (t) |
33,4 |
38,0 |
38,0 |
- |
28,5 |
28,2 |
29,0♂ |
29,0 |
27,0 |
| DT (t) |
21 |
25,0 |
22,0 |
- |
- |
- |
18,5♂ |
17,5 |
17,5 |
| H (t/a) |
23,5/15,8 |
28,0/18,0 |
26,5/18,0 |
- |
21,5/12,7 |
22,0/12,8 |
22,0/14,7♂ |
23,0/15,0 |
19,5/- |
| Metatarsiano III |
| |
LPV-2793 drcho |
395049 drcho |
|
|
- |
|
drcho |
|
|
| DAP (p/m/d) |
28,4/15,7/19,4 |
32,5/17,5/23,0 |
|
- |
22,4/11,1/17,5 |
|
24,5/12,5/18,1♂ |
- |
- |
| DT(p/m/d) |
21,5/16,6/19,7 |
22,0/19,0/22,0 |
|
- |
18,5/15,0/16,0 |
|
18,0/15,8/16,8♂ |
- |
- |
| DT (mx. d) |
21,8 |
22,5 |
|
- |
18,0 |
|
19,6♂ |
- |
- |
| L |
107,8 |
120,0 |
|
99,2 |
86,7 |
|
89,2♂ |
- |
- |
| Yacimientos de procedencia y otro material de comparación: Jaurens (Faure & Guérin, 1983) y Marizulo (Altuna, 1967); más ejemplares actuales de Burgos (1, col. MNCN, Madrid; 3, col. UCM, Madrid), Lyon (Faure & Guérin, 1983) y Basilea (Faure & Guérin, 1983). Abreviaturas en Tabla 1. |
Desde el punto de vista estratigráfico y como bien dedujeron Arsuaga et al. (2010), el material que se analiza en este trabajo, obtenido en la primera fase de excavaciones (1980-1989), procede del nivel estratigráfico
5 de los diferenciados por Pérez-González et al. (2010) en el yacimiento, y en su mayoría deriva de los sectores actualmente denominados como central y su límite con el septentrional.
En las Figs. 1 y 2 reproducimos imágenes de aquella primera fase de excavaciones, incluyendo dos planimetrías del yacimiento en campañas distintas,
antes y después de retirar algunos bloques correspondientes al techo desplomado de la cavidad (Fig. 2). En la cuadrícula A6 es donde según Arsuaga et al. (2012, fig. 3c) se realizó la datación de 90,961 ± 7,761 ka BP para el yacimiento, sobre una muestra no especificada “de sedimento”,
obtenida en un horizonte supuestamente infrayacente a los documentados en nuestra primera fase de excavaciones. Pérez-González et al. (2010, p. 413) y Blain et al. (2014, fig. 2C) mencionan también la existencia de cierto espeleotema asociado a restos de vertebrados, cuya datación por termoluminiscencia
aportó valores aún más antiguos (128,0 ± 12,8 ka / 110,1 ± 11,9 ka). Este nivel se extiende aparentemente entre el sur y centro
del yacimiento, y su litología y datación (MIS 5e, Eemiense) resultaría más próxima a los resultados inéditos con ESR y series
de uranio que en un principio habían sido generalizados para todo el yacimiento (Carbó et al., 1990; Alférez & Roldán, 1992). La datación adoptada en los trabajos pioneros se centró en el espeleotema discontinuo que corona la principal unidad fosilífera,
sellando parcialmente el nivel 5 de los autores posteriores, y que fue muestreado en la cuadrícula A5, donde yace cuarteado
bajo un gran bloque dolomítico desplomado del techo de la cavidad. Este horizonte fue datado en 152 ± 7 ka por ESR y en 118
(+40/-38) ka según los cocientes 230Th/234U y 234U/238U para U= 446 ppb (R. Grün, com. escr., mayo 1985). Finalmente, el espeleotema estalagmítico inferior, excavado esencialmente
en el sector central y meridional del yacimiento, suministró edades absolutas de 206,7 ± 17,7 ka mediante resonancia paramagnética
electrónica (ESR) y 189 (+26/-13) ka mediante series de uranio (230Th/234U y 234U/238U, para U= 256 ppb). El conjunto de estas dataciones, derivadas de muestras tomadas in situ por expertos de la Universidad de Colonia (Alemania) y procesadas en los laboratorios más avanzados de su época (R. Grün,
com. escr., mayo 1985), son las que justificaron con detalles inéditos, la asignación inicial del yacimiento de Pinilla del
Valle al Pleistoceno medio, contribuyendo decisivamente a relacionar los fósiles humanos con formas anteneandertales (Alférez & Roldán, 1992).
|
 Fig. 1—A, Panorámica del yacimiento desde el camino tras la campaña de 1981; B, vista de la cuadrícula A-5 (cota -125 cm) en la campaña
de 1985 con numerosos restos de vertebrados y la mandíbula casi completa de Sus scrofa LPV-2512 in situ (señalada por la flecha); C, detalle de la misma mandíbula en vista lateral izquierda, con un fragmento de asta de cérvido
entre las dos ramas mandibulares. Fig. 1—A, Panorámica del yacimiento desde el camino tras la campaña de 1981; B, vista de la cuadrícula A-5 (cota -125 cm) en la campaña
de 1985 con numerosos restos de vertebrados y la mandíbula casi completa de Sus scrofa LPV-2512 in situ (señalada por la flecha); C, detalle de la misma mandíbula en vista lateral izquierda, con un fragmento de asta de cérvido
entre las dos ramas mandibulares.
|
|
|
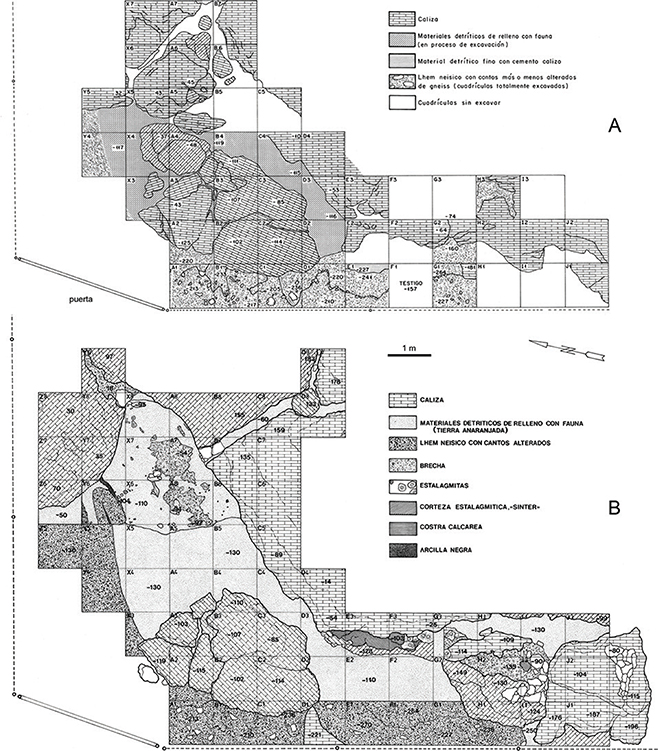 Fig. 2—Esquema en planta de la excavación de la Cueva del Camino a término de las campañas de 1982 (A) y 1988 (B). Se aprecia el
progreso de la excavación hacia el sector septentrional, una vez retirado parte del recubrimiento rocoso derivado del colapso
de la cavidad. Fig. 2—Esquema en planta de la excavación de la Cueva del Camino a término de las campañas de 1982 (A) y 1988 (B). Se aprecia el
progreso de la excavación hacia el sector septentrional, una vez retirado parte del recubrimiento rocoso derivado del colapso
de la cavidad.
|
|
MetodologíaTOP
En el estudio descriptivo de la dentición se ha usado la nomenclatura típica del molar tribosfénico de los Euterios (Alférez, 1990), dando en todo caso las características de algunos dientes típicos de los suidos (por ejemplo, caninos, molares terceros).
En el estudio métrico, la manera de medir los dientes, tomando su longitud y anchura máximos, así como la forma de medir los
diastemas y distancias entre dientes, se ha hecho con arreglo a las normas dadas por Van der Made (1991, 1996).
En todas las tablas donde se dan las dimensiones de los dientes, las piezas que están insertas en restos maxilares o mandibulares,
han sido señaladas con un asterisco (*).
En los restos craneales se han tomado todas aquellas medidas que no han sido afectadas por la rotura o deformación producidas
en el proceso diagenético. Algunas de estas medidas han sido indicadas en los esquemas de estos restos incluidos en el texto
(Figs. 3-4).
|
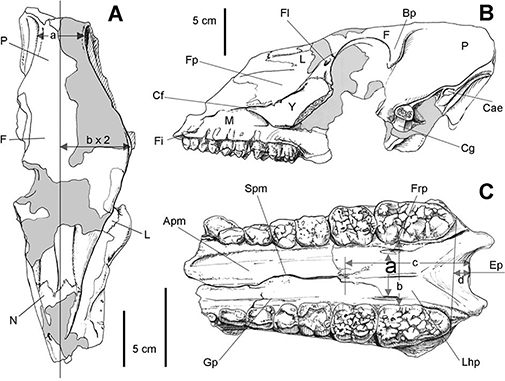 Fig. 3—Sus scrofa, Pleistoceno de Pinilla del Valle (Madrid). A-B, Esquemas del fragmento craneal LPV-1010 en norma dorsal o superior (A) y
lateral izquierda (B); C, esquema en norma ventral o inferior del fragmento craneal LPV-0798 mostrando la dentición yugal.
Abreviaturas: Apm, apófisis palatina del maxilar; Bp, barra postorbitaria; Cae, conducto auditivo externo; Cf, cresta facial;
Cg, cavidad glenoidea; Ep, espina nasal; F, frontal; Fi, foramen infraorbitario; Fl, forámenes lacrimales; Fp, fosa preorbitaria;
Frp, forámenes rostrales del palatino; Gp, garganta palatina; L, lacrimal; Lhp, lámina horizontal del palatino; M, maxilar;
N, nasal; P, parietal; Spm, sutura palatina mediana; Y, yugal, malar o cigomático. Las dimensiones a-d se referencian en las
tablas. Fig. 3—Sus scrofa, Pleistoceno de Pinilla del Valle (Madrid). A-B, Esquemas del fragmento craneal LPV-1010 en norma dorsal o superior (A) y
lateral izquierda (B); C, esquema en norma ventral o inferior del fragmento craneal LPV-0798 mostrando la dentición yugal.
Abreviaturas: Apm, apófisis palatina del maxilar; Bp, barra postorbitaria; Cae, conducto auditivo externo; Cf, cresta facial;
Cg, cavidad glenoidea; Ep, espina nasal; F, frontal; Fi, foramen infraorbitario; Fl, forámenes lacrimales; Fp, fosa preorbitaria;
Frp, forámenes rostrales del palatino; Gp, garganta palatina; L, lacrimal; Lhp, lámina horizontal del palatino; M, maxilar;
N, nasal; P, parietal; Spm, sutura palatina mediana; Y, yugal, malar o cigomático. Las dimensiones a-d se referencian en las
tablas.
|
|
|
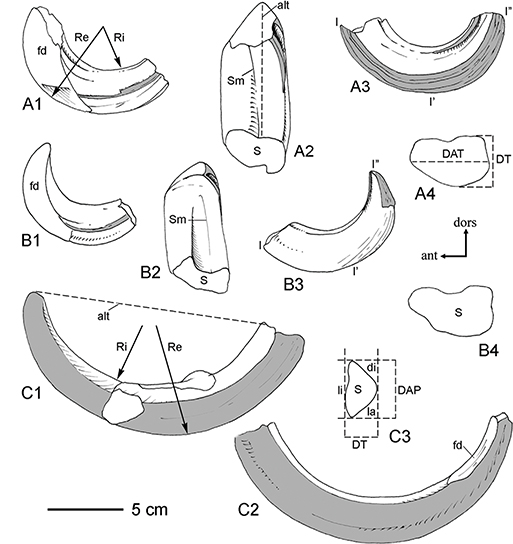 Fig. 4—Sus scrofa, Pleistoceno de Pinilla del Valle (Madrid). Esquemas de algunos elementos dentarios. A-B, caninos superiores derechos LPV-0524
y LPV-2257, respectivamente, representados por su cara anterior (A1, B1), labial (A2, B2) y postero-lingual (A3, B3), más
el contorno de sus secciones posteriores a nivel del cuello (A4, B4). C, canino inferior izquierdo LPV-2209 visto (en C1)
por la cara distal (en blanco) y labial (sombreado); en C2, con la cara lingual sombreada, y la sección posterior correspondiente
a nivel del cuello (C3). Abreviaturas: alt, altura (distancia del ápice al borde labial); DAP, diámetro antero-posterior;
di, cara distal; DT, diámetro transverso; fd, faceta de desgaste; l-l’-l”, longitud del diente siguiendo la línea de máxima
curvatura; la, cara labial; li, cara lingual; Re, Ri, radios de curvatura externo e interno; S, sección en la base (cuello);
Sm, surco mediano. Fig. 4—Sus scrofa, Pleistoceno de Pinilla del Valle (Madrid). Esquemas de algunos elementos dentarios. A-B, caninos superiores derechos LPV-0524
y LPV-2257, respectivamente, representados por su cara anterior (A1, B1), labial (A2, B2) y postero-lingual (A3, B3), más
el contorno de sus secciones posteriores a nivel del cuello (A4, B4). C, canino inferior izquierdo LPV-2209 visto (en C1)
por la cara distal (en blanco) y labial (sombreado); en C2, con la cara lingual sombreada, y la sección posterior correspondiente
a nivel del cuello (C3). Abreviaturas: alt, altura (distancia del ápice al borde labial); DAP, diámetro antero-posterior;
di, cara distal; DT, diámetro transverso; fd, faceta de desgaste; l-l’-l”, longitud del diente siguiendo la línea de máxima
curvatura; la, cara labial; li, cara lingual; Re, Ri, radios de curvatura externo e interno; S, sección en la base (cuello);
Sm, surco mediano.
|
|
En las mandíbulas, además de las series dentarias, se ha medido la longitud de los diastemas, los cuales proporcionan información
sobre la edad y el sexo de un individuo, debido sobre todo a la influencia del tamaño de los caninos en la longitud de estos
diastemas.
En los caninos superiores, a las medias básicas se han añadido otras como la longitud del diente según su mayor curvatura
o el perímetro del mismo en la base. En los caninos inferiores se han tomado las medidas correspondientes a la longitud, altura
y radios de curvatura, junto con las de la sección en la base, de gran valor sistemático.
Todo ello ha sido indicado en los esquemas realizados para estos dientes incluidos en el texto (Fig. 4). Además, este estudio se presenta en forma de tablas comparativas, con abreviaturas normalizadas y dimensiones expresadas
en milímetros. Las medidas menos frecuentes han sido explicadas en los correspondientes textos y figuras.
Por prescripción legal, el material estudiado en este trabajo se integró, junto con el resto de la colección paleontológica
del yacimiento de Pinilla del Valle, en el Museo Arqueológico Regional de la Comunidad de Madrid (Alcalá de Henares), donde
provisionalmente optaron por mantener las siglas “LPV” que designan los fósiles correspondientes a la primera fase de excavaciones
culminada en 1989.
Descripción del materialTOP
Restos cranealesTOP
Se han encontrado dos fragmentos craneales. En uno de ellos (LPV-1010) falta la parte anterior con su dentición, y el otro
fragmento (LPV-0798) sólo conserva la parte media basal, desde el alveolo del canino hasta el borde posterior del palatino.
Ambos restos conservan la dentición yugal casi completa en ambos maxilares. Hay además tres fragmentos de maxilares superiores
con dentición (LPV-1214, LPV-1226; LPV-1413: Tabla 1).
Fragmento craneal LPV-1010 (Figs. 3 y 5)TOP
En norma dorsal o superior (Fig. 3A) puede verse el plano formado por el parietal (P) y el frontal (F) casi completo, aunque no se aprecia ninguna sutura. Los
nasales (N) solo se conservan en su mitad posterior aproximadamente, aunque están deformados por la presión sufrida por el
cráneo.
|
 Fig. 5—Sus scrofa, Pleistoceno de Pinilla del Valle (Madrid). Fragmento craneal LPV-1010 en norma lateral izquierda (A) y ventral (B). Fig. 5—Sus scrofa, Pleistoceno de Pinilla del Valle (Madrid). Fragmento craneal LPV-1010 en norma lateral izquierda (A) y ventral (B).
|
|
En las partes posterior y lateral izquierda, que están bien conservadas, se han tomado las siguientes medidas: anchura mínima
de los parietales (a), 32 mm; anchura máxima de los frontales (b x 2), 98 mm.
En norma ventral o inferior (Fig. 5B) se ve cómo la compresión lateral sufrida por el cráneo afecta a la región palatina, mientras que los maxilares, bien conservados,
portan gran parte de la dentición yugal (Tabla 1). La parte posterior conserva los cóndilos occipitales, con el foramen magnum relleno de sedimento. En esta última región se han tomado las siguientes medidas: distancia entre la parte externa de los
cóndilos, 57 mm; distancia entre los bordes internos de los cóndilos (equivalente al diámetro máximo del foramen magnum), 23 mm.
En norma lateral izquierda (Figs. 3B y 5A) pueden identificarse todos los huesos de esta región no afectada por la rotura del cráneo. En el hueso maxilar (M) se distingue
el foramen infraorbitario (Fi), que aparece roto, situado a la altura de la separación entre el P4 y el M1. Sobre el maxilar (M) y el yugal (Y) aparece la cresta facial (Cf) bien desarrollada y, por encima de ella, la fosa preorbitaria
(Fp), profunda y bien definida. Su pared ósea está hundida y rellena de sedimento.
La región de la órbita presenta una amplia fosa orbitaria abierta en su parte postero-inferior, donde limita con la barra
postorbitaria (Bp). Su borde anterior presenta dos forámenes lacrimales (Fl) a diferencia de los otros suidos, en los que
sólo hay uno. El borde inferior de la órbita, formado por el hueso yugal (Y), malar o cigomático, está curvado hacia atrás,
característica de los suidos modernos. Este hueso cigomático aparece roto (falta prácticamente toda la arcada cigomática),
pero su sección muestra que es un hueso ancho y robusto.
En la parte posterior de la órbita, el parietal (P) y el temporal (T) aparecen fusionados, con unas líneas de fractura que
podrían corresponder a la sutura entre ambos. En la parte inferior del temporal se ve la parte basal de la arcada cigomática;
en su parte inferior está la cavidad glenoidea (Cg), y en la superior, el orificio del conducto o meato auditivo externo (Cae).
En esta región craneal se han tomado las siguientes medidas no afectadas por la deformación: longitud máxima lateral del parietal
(c), 117 mm; diámetro máximo de la órbita (d), 53 mm; anchura del hueso cigomático en su base (e), 44,5 mm.
Fragmento craneal LPV-0798TOP
En este resto craneal (Figs. 3C y 6C) los maxilares con su dentición (Tabla 1) están bien conservados, pero la región del paladar está afectada por la presión sufrida en el proceso diagenético. Así,
vemos que las apófisis palatinas del maxilar (Apm) están fracturadas en su parte media, de modo que la sutura palatina mediana
(Spm) está distorsionada formado una línea discontinua.
|
 Fig. 6—Sus scrofa, Pleistoceno de Pinilla del Valle (Madrid). A, fragmento maxilar derecho LPV-1226 con D2, D3, D4 y M1, caras lingual (A1), labial (A2) y oclusal (A3); B, resto maxilar superior derecho LPV-1214 con P3-M3 en vista oclusal (B1), lingual (B2) y labial (B3); C, fragmento craneal LPV-0798 con maxilares relativamente bien conservados,
mostrando la dentición (P2-M3) en vista oclusal. Fig. 6—Sus scrofa, Pleistoceno de Pinilla del Valle (Madrid). A, fragmento maxilar derecho LPV-1226 con D2, D3, D4 y M1, caras lingual (A1), labial (A2) y oclusal (A3); B, resto maxilar superior derecho LPV-1214 con P3-M3 en vista oclusal (B1), lingual (B2) y labial (B3); C, fragmento craneal LPV-0798 con maxilares relativamente bien conservados,
mostrando la dentición (P2-M3) en vista oclusal.
|
|
La lámina horizontal del palatino (Lhp) tiene forma netamente triangular; en sus suturas con las apófisis palatinas se abren
los forámenes rostrales del palatino (Frp), de forma ovalada y situados a la altura del primer lóbulo del M3. De ellos parten unas gargantas palatinas (Gp) largas y poco marcadas.
En la lámina horizontal del palatino, la distancia (d) entre la línea que une la parte posterior de los M3 con el borde posterior del palatino o espina nasal (Ep) es pequeña, rasgo típico de Sus scrofa.
En este fragmento craneal se han tomado las siguientes medidas no afectadas por la deformación: distancia entre los bordes
externos de los forámenes rostrales del palatino (a), 28,5 mm; anchura palatina tomada entre el primer y segundo lóbulo de
los M3 (b), 38 mm; distancia entre el punto más anterior de la lámina palatina y el borde posterior del palatino (espina nasal)
(c), 76,4 mm; distancia desde la línea que une la parte posterior de los M3 con el borde posterior del palatino (d), 9,7 mm.
Dentición superior (Tabla 1)TOP
Dentición de lecheTOP
Se ha encontrado un fragmento de maxilar juvenil derecho (LPV-1226) con DP2, DP3, DP4, M1, únicos dientes de leche encontrados (Fig. 6A1-A3).
El DP2 es un diente estrecho y alargado parecido al P2, pero de tamaño ligeramente inferior (Tabla 1). Tiene una cúspide central y dos crestas laterales, con el parastilo bien desarrollado. En la parte distal tiene una plataforma
lingual con una pequeña depresión en el centro.
El DP3 tiene una plataforma postero-lingual más desarrollada que el DP2 y además tiene otra más pequeña en la parte anterior, lo que le da un aspecto bilobulado, con la parte anterior más estrecha
que la posterior.
El DP4 tiene un aspecto molariforme muy distinto del P4 definitivo y más parecido al M1. Presenta dos lóbulos con las cuatro cúspides principales y un leve cíngulo en la parte anterior. El lóbulo anterior está
más desplazado hacia labial que en el M1.
IncisivosTOP
I1. ‒ Se ha encontrado un diente derecho completo (Fig. 7G) bastante gastado, con la parte media de la raíz cubierta en parte por sedimento. Es un diente algo curvado con la cara lingual
ligeramente cóncava y la labial suavemente convexa. La superficie oclusal es ovalada, ligeramente cóncava y algo más estrecha
en la parte distal. Está inclinada en sentido antero-posterior (hacia el I2).
|
 Fig. 7—Sus scrofa, Pleistoceno de Pinilla del Valle (Madrid). Elementos de la dentición superior. A, canino superior derecho LPV-0524, caras
lingual (A1) y labial (A2); B, canino superior derecho LPV-2257, caras lateral izquierda (B1), lateral derecha (B3) y labial
(B2); C, P1 derecho LPV-1686, caras lingual (C1) y labial (C2); D, fragmento maxilar izquierdo LPV-1413 con P2 y P3 en vista lingual (D1), labial (D2) y oclusal (D3); E, P3 derecho LPV-0321 en vistas lingual (E1), oclusal (E2) y labial (E3); F, P3 izquierdo LPV-0430, caras lingual (F1), oclusal (F2) y labial (F3); G, M3 derecho LPV-0320, caras oclusal (H1) y labial (H2). Escalas gráficas 10 mm, excepto A, B y D (20 mm). Fig. 7—Sus scrofa, Pleistoceno de Pinilla del Valle (Madrid). Elementos de la dentición superior. A, canino superior derecho LPV-0524, caras
lingual (A1) y labial (A2); B, canino superior derecho LPV-2257, caras lateral izquierda (B1), lateral derecha (B3) y labial
(B2); C, P1 derecho LPV-1686, caras lingual (C1) y labial (C2); D, fragmento maxilar izquierdo LPV-1413 con P2 y P3 en vista lingual (D1), labial (D2) y oclusal (D3); E, P3 derecho LPV-0321 en vistas lingual (E1), oclusal (E2) y labial (E3); F, P3 izquierdo LPV-0430, caras lingual (F1), oclusal (F2) y labial (F3); G, M3 derecho LPV-0320, caras oclusal (H1) y labial (H2). Escalas gráficas 10 mm, excepto A, B y D (20 mm).
|
|
I2 ‒ Sólo se ha encontrado un ejemplar completo perfectamente conservado (Fig. 9H). Es un diente aplanado buco-lingualmente y de aspecto subtriangular debido a que su única raíz se estrecha gradualmente
hacia el ápice y se incurva ligeramente hacia mesial. En la corona, la línea cervical es más o menos paralela al borde oclusal,
y en su parte media hay una ligera depresión que delimita una parte posterior inclinada suavemente hacia lingual. La superficie
oclusal, poco gastada, es estrecha y alargada, con una faceta de desgaste ovalada en su parte anterior.
|
 Fig. 8—Sus scrofa, Pleistoceno de Pinilla del Valle (Madrid). Restos mandibulares LPV-0315 (A) y LPV-2512 (B), con los caninos y la dentición
yugal casi completa. El ejemplar B se presenta fracturado por la carga sedimentaria, pero conserva bien los incisivos en la
región sinfisaria, a pesar de haber sufrido daños en su depósito en el museo (comparar Fig. 1B-C). Fig. 8—Sus scrofa, Pleistoceno de Pinilla del Valle (Madrid). Restos mandibulares LPV-0315 (A) y LPV-2512 (B), con los caninos y la dentición
yugal casi completa. El ejemplar B se presenta fracturado por la carga sedimentaria, pero conserva bien los incisivos en la
región sinfisaria, a pesar de haber sufrido daños en su depósito en el museo (comparar Fig. 1B-C).
|
|
|
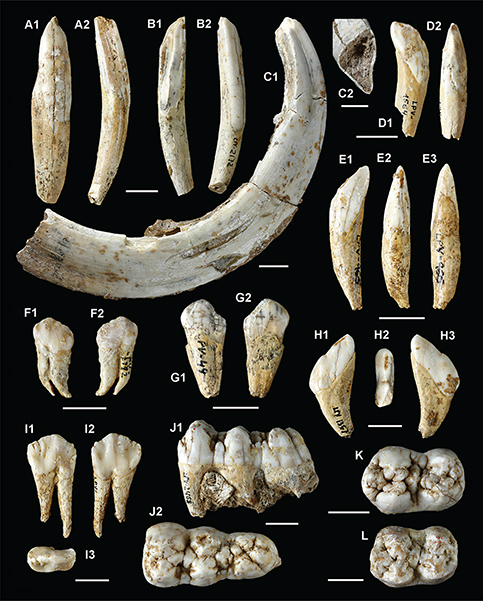 Fig. 9—Sus scrofa, Pleistoceno de Pinilla del Valle (Madrid). Elementos de la dentición inferior (excepto H). A, I2 derecho LPV-1383, caras lateral (A1) y oclusal (A2); B, I2 izquierdo LPV-2172, caras oclusal (B1) y lateral (B2); C, canino inferior izquierdo LPV-2209 visto por la cara lingual (C1),
y sección posterior nivel del cuello (C2); D, I3 izquierdo LPV-1564, caras labial (D1) y oclusal (D2); E, I3 izquierdo LPV-0966, caras lingual (E1), oclusal (E2) y labial (E3); F, P1 derecho LPV-2792, caras lingual (F1) y labial (F2); G, P1 izquierdo, caras labial (G1) y lingual (G2); H, I2 derecho LPV-1357, caras lingual (H1), oclusal (H2) y labial (H3); I, P3 derecho LPV-1178, caras labial (I1), lingual (I2) y oclusal (I3); J, M3 izquierdo LPV-2468, caras labial (J1) y lingual (J2); K, M2 izquierdo LPV-1228 en vista oclusal; L, M2 izquierdo, LPV-1534, cara oclusal. Escalas gráficas, 10 mm Fig. 9—Sus scrofa, Pleistoceno de Pinilla del Valle (Madrid). Elementos de la dentición inferior (excepto H). A, I2 derecho LPV-1383, caras lateral (A1) y oclusal (A2); B, I2 izquierdo LPV-2172, caras oclusal (B1) y lateral (B2); C, canino inferior izquierdo LPV-2209 visto por la cara lingual (C1),
y sección posterior nivel del cuello (C2); D, I3 izquierdo LPV-1564, caras labial (D1) y oclusal (D2); E, I3 izquierdo LPV-0966, caras lingual (E1), oclusal (E2) y labial (E3); F, P1 derecho LPV-2792, caras lingual (F1) y labial (F2); G, P1 izquierdo, caras labial (G1) y lingual (G2); H, I2 derecho LPV-1357, caras lingual (H1), oclusal (H2) y labial (H3); I, P3 derecho LPV-1178, caras labial (I1), lingual (I2) y oclusal (I3); J, M3 izquierdo LPV-2468, caras labial (J1) y lingual (J2); K, M2 izquierdo LPV-1228 en vista oclusal; L, M2 izquierdo, LPV-1534, cara oclusal. Escalas gráficas, 10 mm
|
|
CaninosTOP
Tenemos dos ejemplares derechos (Figs. 4A-B y 7A-B), uno de ellos con el extremo apical roto, y un fragmento de la cara lingual de otro izquierdo. Son dientes gruesos y robustos,
con una curvatura ligeramente espiralada, dirigida hacia arriba.
La cara lingual, convexa, está formada por tres franjas o bandas de esmalte: una anterior con pocas estrías; otra ventral,
ancha y con numerosas estrías o acanaladuras paralelas a la curvatura del diente; y una tercera posterior, lisa y con una
ligera depresión central. Las dos primeras terminan en la faceta de desgaste.
La cara labial, cóncava, es ancha, más o menos lisa, y está dividida en dos lóbulos por un surco mediano poco profundo, más
ancho en la parte distal.
En estos dientes se ha tomado el radio de curvatura, tanto externo como interno, considerado como el radio de la circunferencia
que contiene la máxima curvatura del diente.
PremolaresTOP
Tenemos 17 premolares; de ellos 13 están implantados en fragmentos craneales y maxilares (Tabla 1) y 4 se han encontrado sueltos. La serie premolar aumenta en tamaño y complicación del P1 al P4, y todos presentan una cúspide principal. Del P2 al P4 tienen una plataforma postero-lingual excavada en el centro, y otra antero-lingual de menor tamaño. Estas plataformas y los
estilos van desarrollándose más del primero al último.
P1 (Fig. 7C). ‒ Se han encontrado dos ejemplares derechos sueltos completos y bien conservados (Tabla 1). Es un diente más bien aplanado, con dos raíces casi unidas y ligeramente arqueadas hacia distal. La corona es baja y con
la cúspide central poco desarrollada. En su cara labial tiene dos surcos (el posterior poco marcado) que delimitan las cúspides
anterior y posterior. La cara oclusal es alargada y engloba las tres cúspides.
P2 (Figs. 6C y 7D). ‒ Tenemos cuatro ejemplares insertos en fragmentos mandibulares (Tabla 1). Es un diente con la corona estrecha y alargada, de aspecto subtriangular. Las plataformas linguales están muy poco desarrolladas,
y los estilos apenas marcados. La superficie oclusal labial tiene una faceta anterior alargada y otra posterior más corta.
P3 (Figs. 5, 6B-C y 7D-F). ‒ Disponemos de siete ejemplares, de los que cinco están en fragmentos maxilares y dos sueltos (Tabla 1) Son dientes bastante más anchos que el P2 debido al desarrollo de la plataforma lingual anterior y, sobre todo, la posterior, lo que le da un aspecto más cuadrangular.
Tienen dos raíces robustas que divergen unos 30˚. Los estilos están bien desarrollados, con el parastilo algo más prominente
que el metastilo. La superficie oclusal labial es recta, alargada, y contiene la cúspide central y las facetas anterior y
posterior.
P4 (Figs. 5 y 6A, C). ‒ Hay cinco ejemplares, todos procedentes de fragmentos maxilares (Tabla 1). Se trata de un diente de aspecto cuadrangular, con las cuatro cúspides principales separadas por surcos profundos que convergen
en el centro. El paracono y el metacono están unidos. El protocono está formado por pequeños tubérculos, y el hipocono está
bien desarrollado. Los estilos están poco marcados.
MolaresTOP
Tenemos 17 molares; de ellos, 15 están implantados en fragmentos craneales y maxilares (Tabla 1) y 2 se han encontrado sueltos. Tienen una morfología parecida, con dos lóbulos transversales al eje longitudinal del diente,
formados por las cuatro cúspides principales (protocono, paracono, metacono, hipocono). Ambos lóbulos están separados por
un valle con una cúspide central de menor tamaño que las principales. Tienen un fuerte cíngulo mesial y otro distal. El M3 tiene además un tercer lóbulo posterior subtriangular, separado también por un valle.
M1 (Figs. 5 y 6A-C). ‒ Hay 7 ejemplares, seis en sus maxilares y uno suelto (Tabla 1). Son dientes rectangulares, con la mitad anterior ligeramente más estrecha que la posterior y el cíngulo mesial algo más
alargado que el distal. El valle es estrecho y está cerrado en su parte lingual por un pequeño tubérculo procedente del hipocono.
M2 (Figs. 5 y 6A, C). ‒ Tenemos cinco dientes insertos en fragmentos maxilares (Tabla 1). Son dientes rectangulares, con los cíngulos anterior y posterior igualmente desarrollados. La cúspide central del valle
está más unida al lóbulo posterior que al anterior. El paracono sobresale ligeramente hacia la cara labial. El valle central
está cerrado en su parte labial por un pequeño tubérculo procedente del metacono.
M3 (Figs, 5, 6A y C, 7H). ‒ Tenemos seis ejemplares, cinco en fragmentos maxilares (Tabla 1) y otro suelto. Es un diente característico, con un tercer lóbulo o talón de forma subtriangular, y con un fuerte cíngulo
mesial. Las dos cúspides labiales (paracono y metacono) están desplazadas hacia delante con relación a las linguales (protocono
e hipocono). El valle entre los dos lóbulos anteriores es estrecho, ocupado por la cúspide ventral y otras accesorias y, con
el uso, solo queda abierto por su parte labial. El tercer lóbulo o talón está formado por 4 o 5 tubérculos o cúspides principales
y alguna otra accesoria. El pequeño valle anterior desaparece con el uso.
Restos mandibularesTOP
Se han encontrado tres mandíbulas incompletas, cada una de ellas con las dos hemimandíbulas unidas por la sínfisis (Fig. 8). En las tres faltan casi por completo las ramas verticales ascendentes, y dos de ellas están algo deformadas debido a la
carga sedimentaria.
Los incisivos solo existen en una de ellas, mientras que las tres conservan los caninos y la dentición yugal casi completa.
Se ha encontrado además un fragmento mandibular izquierdo con P4-M3, así como un resto aislado correspondiente a un cóndilo mandibular. En estos restos mandibulares se han tomado las siguientes
medidas, aparte de las citadas para la dentición:
| • |
Segmentos dentarios en las series yugales existentes (Tabla 4) |
| • |
Longitud de los diastemas y distancias entre los caninos y los premolares. |
| • |
Anchura mandibular en la región sinfisaria (Tabla 4) |
Cóndilo mandibularTOP
Es un fragmento de la rama ascendente de una mandíbula, que conserva completo el proceso articular y la parte superior de
dicha rama, entre la escotadura supracondilea y el borde posterior de la mandíbula.
El cóndilo es alargado transversalmente y de forma subcilíndrica, con la parte externa más gruesa y corta que la parte interna,
que es más apuntada. La distancia entre sus extremos es 23,2 mm.
La anchura entre la base anterior del cóndilo y el borde posterior de la mandíbula es 33,5 mm.
Dentición inferior (Tabla 2)TOP
Dentición de lecheTOP
Sólo se ha encontrado un DI1 (Tabla 2). Es un diente casi rectilíneo, ligeramente curvado hacia mesial en la parte superior de la corona. Su tamaño es bastante
menor que el I1 definitivo.
IncisivosTOP
Hay cinco incisivos mal conservados en la mandíbula LPV-2512 y otros siete sueltos: 3 I2 y 4 I3.
En los dientes existentes en esta mandíbula (Fig. 8B) puede verse que están implantados casi horizontalmente, con los I2 convergiendo hacia la sínfisis. Los I3 están algo separados, y los I1 son bastante menores que los I2. Estos últimos son dientes muy alargados (la longitud media es de unos 65 a 70 mm) y ligeramente curvados, con la superficie
oclusal algo inclinada hacia distal.
Los I3 son bastante más cortos (32 mm de media) y ligeramente curvados. La corona, igual que en los otros incisivos, es alargada,
y en la cara labial alcanza la mitad del diente. La superficie oclusal es crenulada con dos pequeñas cúspides anteriores.
Tiene una ligera inclinación hacia distal que se va acentuando con el desgaste.
CaninosTOP
Tenemos siete ejemplares, de los que seis están en los restos mandibulares y otro suelto (Tabla 3). Este último está bastante bien conservado (Figs. 4C y 9C), aunque con una ligera rotura (3 o 4 mm) en la parte apical. Cinco de ellos corresponden a individuos machos, como indica
su gran tamaño, y los otros dos, bastante más pequeños, corresponden a un resto mandibular de hembra (LPV-1009).
Existe además un resto de otro canino derecho que conserva casi toda la cara lingual con el ápice roto. Las únicas medidas
con un grado de aproximación fiable que han podido tomarse (cara lingual: aproximadamente 25 mm; altura: aproximadamente 110
mm; Re=69; Ri=58) así como su aspecto y morfología, lo asemejan extraordinariamente al canino LPV-2209, por lo que es probable
que ambas defensas pertenecieran al mismo individuo.
Los caninos inferiores son dientes alargados, curvados hacia atrás de forma ligeramente espiralada y aspecto semicircular.
La sección de la base (a nivel alveolar) forma un triángulo asimétrico con los caracteres típicos del llamado “tipo scrofa o scrofiano”, con el lado labial (antero-externo) más corto que el lingual.
Las caras labial y lingual están en parte recubiertas de esmalte, siendo esta superficie esmaltada más amplia en la cara lingual
que en la labial.
En la cara lingual (antero-interna) hay un surco poco profundo que sigue la curvatura del diente y aparece situado algo más
próximo al borde mesial que al distal.
Disponemos de 24 premolares (Tabla 2), de los que 21 están insertos en los fragmentos mandibulares (Tabla 4) y los otros tres fueron encontrados aislados.
Los cuatro premolares tienen una estructura análoga. Son dientes birradiculados, con la raíz posterior más robusta que la
anterior. En los dientes poco usados, la corona es subtriangular, con el borde crenulado. Con el desgaste aparecen tres cúspides:
una central (doble) y otras dos correspondientes a los estílidos anterior y posterior. A la vez, la corona se hace ondulada,
con la parte anterior más alta y la posterior más baja y convexa. El estílido posterior y su cúspide están más desarrollados
que el anterior.
Disponemos de 24 molares inferiores (Tabla 2), de ellos 21 se encuentran en los fragmentos mandibulares (Tabla 4) y 3 se han encontrado sueltos.
Están formados básicamente por dos lóbulos con las cúspides principales (protocónido, paracónido, metacónido e hipocónido),
separados por un valle transversal al eje longitudinal del diente, en cuyo centro hay una colina o tubérculo de menor tamaño
que las principales. El M3 posee además un tercer lóbulo o talónido, que le da un aspecto subtriangular.
M1 (Figs. 8A-B). ‒ Hay 7 ejemplares implantados en los fragmentos mandibulares existentes (Tablas 2 y 4).
M2 (Figs. 9K-L). ‒ Hay 9 ejemplares; 7 en los fragmentos mandibulares y 2 sueltos (Tablas 2 y 4).
Estos dos dientes responden a un mismo modelo. Ambos tienen forma rectangular, con el lóbulo anterior algo más estrecho que
el posterior. En el valle central tienen un tubérculo que lo cierra labialmente. En la cara anterior hay un pequeño cíngulo,
y en la parte posterior central hay una cúspide o hipoconúlido que acaba uniéndose al cíngulo distal.
M3 (Figs. 8A-B y 9J). ‒ Tenemos 8 ejemplares sin desgastar o poco gastados, 7 de ellos insertos en los fragmentos mandibulares y otro suelto
sin desgastar (Tablas 2 y 4). Existe además un germen (LPV-0795) al que le falta el talónido y cuyo aspecto indica que ha sido atacado por ácidos, es
decir, digerido por un carnívoro.
Son dientes alargados, subtriangulares, con un cíngulo mesial. Están formados por tres lóbulos: dos anteriores con los dos
pares de cúspides principales, y un tercer lóbulo o talónido que tiene otro par de cúspides anteriores, algo más pequeñas
que las principales, y otras tres (o más) menores, que forman la parte final del diente. La anchura de los lóbulos va disminuyendo
del primero al último (Tabla 2).
En los dos valles que separan los lóbulos, además de los tubérculos centrales hay otros más pequeños que los cierran labialmente.
|
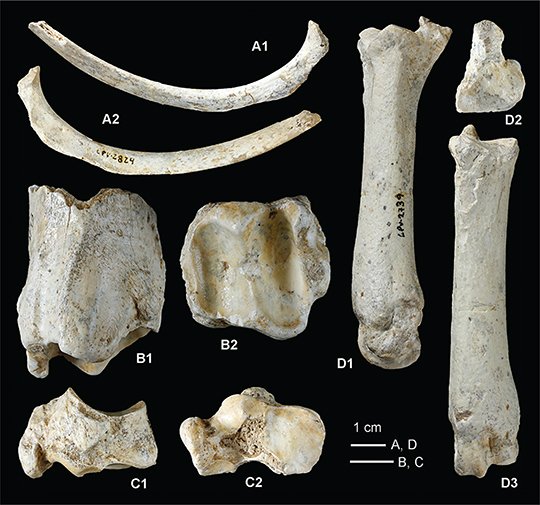 Fig. 10—Sus scrofa, Pleistoceno de Pinilla del Valle (Madrid). Elementos postcraneales. A, Costilla LPV-2824 en vista ventral (A1) y dorsal
(A2); B, Fragmento distal derecho de la tibia LPV-1743 en vista posterior (B1), y cara inferior de articulación con el astrágalo
(B2); C, navicular izquierdo LPV-1507 en vista lateral o externa (C1) y cara distal o inferior (C2); D, metatarsiano III derecho
LPV-2739 visto por su cara externa (D1), desde la cara articular proximal (D2) y por la cara anterior (D3). Fig. 10—Sus scrofa, Pleistoceno de Pinilla del Valle (Madrid). Elementos postcraneales. A, Costilla LPV-2824 en vista ventral (A1) y dorsal
(A2); B, Fragmento distal derecho de la tibia LPV-1743 en vista posterior (B1), y cara inferior de articulación con el astrágalo
(B2); C, navicular izquierdo LPV-1507 en vista lateral o externa (C1) y cara distal o inferior (C2); D, metatarsiano III derecho
LPV-2739 visto por su cara externa (D1), desde la cara articular proximal (D2) y por la cara anterior (D3).
|
|
Tenemos una costilla rota en su extremidad distal a la altura de los cartílagos costales. La región proximal conserva la cabeza
y el tubérculo para la articulación con la vértebra correspondiente. El ángulo distal, en la zona de curvatura, está a unos
18 mm del tubérculo. La longitud total del fragmento conservado es 95 mm.
Hay un fragmento distal derecho roto un poco por encima de la región supraarticular. Longitud: 5 cm.
La cara inferior, de articulación con el astrágalo, está formada por dos gargantas paralelas separadas por un relieve prominente.
La garganta interna es más profunda, estrecha y alargada que la externa. El relieve intermedio entre ambas poleas es cóncavo
en sentido antero-posterior.
En la cara anterior, las apófisis o salientes correspondientes al relieve intermedio y al maléolo interno están prácticamente
a la misma altura. Entre estas dos apófisis hay una zona de tuberosidad.
En la cara externa, la superficie articular para la fíbula o peroné es de pequeño tamaño, ligeramente cóncava y con el borde
superior sinuoso. Junto a ella, la zona de inserción muscular es amplia y bien marcada.
En la cara postero-interna, el surco para el tendón flexor es ancho, poco profundo y menos marcado que en los rumiantes.
Tenemos un ejemplar izquierdo muy bien conservado. Es un hueso alargado en sentido antero-posterior y más bien estrecho.
La cara proximal o superior está formada por dos gargantas para la articulación con el astrágalo, dirigidas antero-posteriormente
y separadas por un relieve cóncavo. La garganta externa es más ancha que la interna, la cual está muy inclinada hacia la parte
medial.
La cara medial o interna tiene tres facetas para la articulación con el cuboides: dos anteriores, pequeñas, estrechas y alargadas,
y otra posterior, de mayor tamaño, cóncavo-convexa, y contorno subcuadrangular.
La cara distal o inferior tiene tres facetas: una anterior para la articulación con el tercer cuneiforme (gran cuneiforme
o ectocuneiforme), ligeramente ondulada y forma cuadrangular, que ocupa la mitad anterior de esta cara. A continuación, en
la parte externa, hay otras dos facetas separadas por pequeños relieves. La primera, pequeña, para el segundo cuneiforme (mesocuneiforme),
y la otra, de tamaño intermedio, para el primer cuneiforme (entocuneiforme).
La cara anterior es cuadrada, con el borde superior recto, y el inferior ligeramente ondulado.
La cara lateral o externa tiene en su parte postero-distal una apófisis ancha y redondeada dirigida hacia abajo que, junto
con la tuberosidad existente en esta cara, sirve como zona de inserción muscular.
Tenemos un solo ejemplar derecho bien conservado.
La epífisis proximal tiene en su centro un pequeño saliente en forma de pico. La parte anterior de la cara articular proximal
es ancha y porta dos facetas articulares: la interna, amplia y cóncavo-convexa, para el tercer cuneiforme; y la externa, pequeña,
ovalada e inclinada lateralmente, para el segundo cuneiforme. La parte posterior está formada por una apófisis o saliente,
dirigida hacia atrás, en forma de lámina más bien estrecha. La cara articular muestra un contorno subtriangular.
La cara anterior es de aspecto rectangular, con el borde interno prácticamente rectilíneo y el externo ligeramente cóncavo.
La cara posterior tiene en la parte proximal la cara posterior de la apófisis, con una faceta ovalada para el sesamoideo.
En la cara externa hay dos pequeñas facetas proximales, algo erosionadas, para el Mt II.
La cara interna es muy aplanada y de aspecto rectangular. Tiene tres facetas proximales para el Mt IV.
En la epífisis distal, el cóndilo interno es más saliente que el externo, formando una tróclea asimétrica para la primera
falange correspondiente.
Dimensiones comparadas de los restos de Sus scrofa de Pinilla del ValleTOP
Las dimensiones de los restos de Sus scrofa de Pinilla del Valle (Tablas 1-4, 6) se han comparado con las medidas de los restos homólogos de otros yacimientos cuaternarios europeos y españoles. Sin embargo,
dado que los restos de jabalíes son, generalmente, muy escasos en los yacimientos, estas medidas tampoco son frecuentes en
la bibliografía utilizada.
Entre los yacimientos europeos con una abundancia relativa de restos de suidos, destacan algunos franceses como Fontèchevade,
La Ferrasie y, sobre todo, el de la cueva de Jaurens (Würm superior) en el suroeste de Francia (Faure & Guérin, 1983) donde, además de las medidas de los restos del yacimiento, se dan muchos datos de otros restos fósiles y actuales existentes
en varios museos europeos.
En las tablas comparativas también se han introducido las dimensiones de ejemplares actuales pertenecientes al Museo Nacional
de Ciencias Naturales (MNCN) de Madrid, y a las colecciones docentes y de comparación de las Facultades de Biología y Geología
de la Universidad Complutense de Madrid (UCM), lo que también ha permitido la comparación de nuestros restos con formas actuales
españolas.
Discusión y resultadosTOP
Identificación taxonómicaTOP
El estudio llevado a cabo sobre la morfología y biometría de la dentición y, sobre todo, de algunos rasgos típicos presentes
en los caninos inferiores, molares terceros y en el cráneo, permiten clasificar inequívocamente estos restos como pertenecientes
a la especie Sus scrofa L., afirmando la clasificación provisional dada en Alférez et al. (1982).
Entre estos rasgos típicos de Sus scrofa que presenta nuestro material, podemos destacar los siguientes:
En la dentición:
| - |
Los últimos molares (M3), tanto superiores como inferiores, presentan el tercer lóbulo (talón y talónido respectivamente)
bien desarrollados y complicados por la presencia de numerosas cúspides (4 a 6 o más). Además, en los M3, entre este tercer lóbulo y las cúspides principales del segundo lóbulo, existe otra cúspide central y varias más pequeñas. |
| - |
Todos los caninos inferiores tienen una sección transversal de forma triangular típica (tipo “scrofa”) con la cara lingual mayor que la labial. |
En el cráneo:
| - |
La región orbital es amplia y presenta dos forámenes lacrimales, a diferencia de otras especies, donde solo existe uno. |
| - |
El borde inferior de la órbita, formado por el hueso yugal, está curvado hacia atrás, característica de los suidos modernos. |
| - |
En la región palatina, la distancia desde la línea que une la parte posterior de los M3 hasta el borde posterior del palatino (espina nasal), es pequeña en relación a las restantes especies de Sus. |
En lo que respecta a la subespecie, los restos de Pinilla se distinguen claramente de Sus scrofa mosbachensis Kuthe por la estructura de los molares terceros, así como por unas dimensiones mucho más reducidas en su conjunto. Se distingue
también de Sus scrofa priscus Goldfuss, por la estructura de los M3, con un talón simple, muy diferente de la descrita anteriormente para nuestros restos, y por los M3, con un talónido con dos colinas poco marcadas en la parte anterior, seguidas de una colina terminal reducida, también muy
diferente de los nuestros.
En cambio, todos los caracteres obtenidos en nuestros restos coinciden con los propios de Sus scrofa scrofa L., dentro de su variabilidad, por lo que los incluimos en esta subespecie.
Dimorfismo sexual y altura en la cruzTOP
El conjunto de los restos de Sus scrofa extraídos del yacimiento, corresponden, según el método empleado por Altuna (1980) a un mínimo de seis individuos.
En este material, el canino superior LPV-0524 (Tablas 3 y 6), robusto y de grandes dimensiones, corresponde a un macho de gran talla y adulto, como indica su desgaste.
De los tres restos mandibulares, dos pertenecen a individuos machos como se deduce del gran tamaño de sus caninos, y el otro,
con estos dientes bastante más pequeños, corresponde a una hembra (Tabla 3). En los dos individuos machos, uno de ellos presenta el M3 sin apenas desgaste y el otro con poco desgaste. Además, hay un M3 aislado (LPV-2468) completamente sin desgastar. Esto indica la presencia de individuos de entre dos años (en los que no hay
desgaste) y tres años de edad (con algo de desgaste) (Mariezkurrena, 1983). La dentición de la hembra, con un desgaste medio, indica un animal adulto de más de tres años.
Tenemos además un fragmento de maxilar (LPV-1226) con la dentición de leche y el M1 casi sin desgastar, que corresponde a un jabato de menos de un año de edad.
Así pues, tenemos un mínimo de seis individuos, entre los que se encuentran un macho adulto, dos machos adolescentes, dos
o tres hembras de distinta edad y un jabato de menos de un año, lo que corresponde a una asociación de individuos típica en
los jabalíes.
Los restos postcraneales, al ser muy escasos, no proporcionan datos estadísticos seguros. De todos modos, en las dimensiones
comparadas (Tabla 5) podemos ver que son algo menores que los de Jaurens, pero sobrepasan a los restos homólogos de las formas actuales tanto
de España como de las europeas de las que tenemos datos.
Al único metatarsiano III hallado, le hemos aplicado el índice de Teichert (1969), dando como resultado una altura en la cruz de 100 cm. Este indica un mayor tamaño de este individuo con respecto a las poblaciones
modernas de esta subespecie, aunque no permite una atribución sexual.
Nuestros resultados para S. scrofa superan claramente a los obtenidos en la segunda fase de excavaciones en el yacimiento, para los que se habían establecido
índices NSPI (nº de ejemplares identificables) = 24 y NMI (nº mínimo de individuos) = 2, representando en torno al 2% de la
asociación de ungulados (Arsuaga et al., 2010; Álvarez-Lao et al., 2013).
Deducciones comparativasTOP
El examen de las tablas de comparación proporciona algunos daros resaltables.
En primer lugar hay que destacar la similitud de los restos dentarios de Pinilla del Valle con los de Jaurens (Pleistoceno
superior; Würm), Mosbach y Mauer (Pleistoceno medio bajo), La Fage (Riss) y Süssernborg (Pleistoceno medio bajo). Concretamente,
los caninos y molares superiores, así como la dentición yugal inferior (Tabla 5) tienen unas medidas prácticamente coincidentes, con diferencias (cuando las hay) que oscilan en general entre uno y dos
milímetros. Los dos caninos inferiores de Pinilla del Valle y Jaurens también son prácticamente iguales (Tabla 5), mientras que los de Mosbach son algo menores.
Esto pone de relieve que en, al menos, cinco sitios europeos de latitudes muy distintas y pertenecientes a diferentes periodos
del Cuaternario, existieron poblaciones de jabalíes de una talla totalmente equiparable dentro de una misma subespecie.
En cuanto a los restos fósiles de finales del Pleistoceno descritos por Altuna (1967, 1973) y Altuna & Mariezkurrena (1984) en yacimientos y cuevas del Cantábrico y del País Vasco, las medidas disponibles (Tablas 5 y 7) también son muy similares a las mencionadas en el párrafo anterior y asimilables, por tanto, a dichas poblaciones. Los datos
expuestos anteriormente parecen no concordar con la hipótesis de una variación de talla en las poblaciones de Sus según pertenezcan a periodos glaciares o interglaciares (Faure & Guérin, 1983).
Respecto a las formas actuales, al comparar las medidas de jabalíes españoles (colecciones de Burgos y El Pardo) con las de
otras poblaciones europeas (Tablas 5 y 7) vemos que hay una coincidencia notable entre ellas, mostrando que, al menos, las poblaciones de jabalíes actuales representadas
por estos datos, tienen tallas prácticamente idénticas.
Consideraciones paleoecológicasTOP
Los restos relativamente abundantes de suidos encontrados en el yacimiento, proporcionan interesantes datos paleoclimáticos,
que confirman y completan la información sobre los ambientes y biotopos que existían en la región, obtenida del conjunto de
la fauna.
El jabalí es un animal de hábitat templado/cálido considerado como un componente típico de las “faunas cálidas” del Cuaternario
(Bonifay, 1966), eminentemente forestal, en bosques mixtos más o menos abiertos, pero con zonas espesas de zarzas y matorral donde encuentra
refugio. También precisa de zonas húmedas, lagunares o de corrientes fluviales, con orillas o charcas más o menos cenagosas,
frecuentando también a veces espacios abiertos de pradera o llanura arbolada.
La existencia de estos hábitats viene confirmada por la información obtenida de otros taxones presentes en el yacimiento.
Asi, entre la fauna de roedores (Toñi & Molero, 1990; Arsuaga et al., 2012; Laplana et al., 2013; Laplana, 2014; Laplana & Sevilla, 2016), las especies Sciurus vulgaris, Elyomis quercinus y Myodes glareolus, son propias de zonas forestales templadas (bosque caducifolio), mientras que la mayor parte de los topillos, como Microtus arvalis, Microtus agrestis y el hámster (Allocricetus bursae) prefieren zonas más descubiertas. Algunos topillos (Microtus gr. duodecimcostatus) y el topo (género Talpa) habitan las praderas húmedas, en tanto que la presencia de musarañas como Crocidura russula indica praderas secas.
Otros roedores presentes en el yacimiento, como Arvicola sapidus, Castor fiber y el mustélido Lutra lutra, además de las aves Anátidas y peces (Salmo sp.) son exponentes de un biotopo ripícola con embalsamientos acuáticos.
Dentro de los macromamíferos, la mayoría de los artiodáctilos identificados (Cervus elaphus, Dama geiselana, Capreolus capreolus) junto con Vulpes vulpes, Canis lupus y el úrsido, también confirman la existencia de bosque caducifolio. A su vez, la presencia del uro (Bos primigenius) junto con los grandes carnívoros (Panthera leo, Panthera? sp., Crocuta crocuta), caballos (Equus caballus pinillensis -nomen nudum- según Alférez et al., 1985; E. caballus torralbae según Maldonado, 1996; E. ferus según Arsuaga et al., 2012; E. caballus según Álvarez-Lao et al., 2013) y rinocerontes (Stephanorhinus hemitoechus), denotan la existencia de estepas próximas a cursos fluviales de aguas permanentes. Por último, la especie Rupicapra rupicapra habita los roquedales de alta montaña.
Por lo expuesto anteriormente, así como por las características geográficas actuales del Valle del Lozoya se deduce, para
la época en que se formó el yacimiento, la presencia de un río en la base del valle rodeado por estepas y distintos tipos
de praderas, mientras que los bosques templados caducifolios se situarían en las laderas de los montes, separando las estepas
basales de los roquedales de alta montaña (cuyas condiciones climáticas serían más frías y rigurosas).
Nuestros resultados paleoecológicos coinciden, pues, con la reconstrucción paleoambiental presentada por Arsuaga et al. (2010, 2012), que además combina los resultados del análisis polínico y antracológico de los sedimentos acumulados en la Cueva del Camino
(Ruiz Zapata et al., 2012), con diversas consideraciones tafonómicas obtenidas en la segunda fase de excavaciones en la misma. La integración con los
datos derivados de las asociaciones de anfibios y reptiles identificados en la Cueva del Camino (Blain et al., 2014), ha proporcionado recientemente un marco mucho más preciso para entender el impacto ecológico de las fluctuaciones climáticas
registradas entre el MIS 5 y los comienzos de MIS 4 en los distintos niveles del yacimiento.
AGRADECIMIENTOSTOP
A la Dra. Maeva Orliac (Instituto de Ciencias de la Evolución, Universidad de Montpellier, Francia) por los excelentes dibujos
de parte del material, realizados durante una visita predoctoral en Madrid. Al prof. Rainer Grün (Centro de Investigación
Australiano en Evolución Humana, Nathan, Australia), por sus primeras dataciones inéditas del yacimiento. A los revisores
científicos Dres. Alfonso Arribas (Museo Geominero, Madrid) y Juan Luis Arsuaga (Universidad Complutense de Madrid) por sus
comentarios y sugerencias, que han permitido mejorar y actualizar el manuscrito. A Carlos Alonso (Universidad Complutense
de Madrid) por las fotografías y el montaje de las ilustraciones. El Dr. César Laplana (Museo Arqueológico Regional, Alcalá
de Henares) atendió gentilmente los requerimientos sobre el material de S. scrofa obtenido en nuestra primera fase de excavaciones, y depositado previamente en el Museo, sobre el que Mario Torquemada tomó
las fotografías reunidas en las figuras 5 y 8. Finalmente, los autores queremos expresar un agradecimiento muy especial al Dr. Juan Carlos Gutiérrez Marco (Instituto de
Geociencias CSIC-UCM, Madrid), por sus ánimos continuados y la ayuda decisiva para la publicación del trabajo.
ReferenciasTOP
| ○ |
Alférez, F. (1985). Dos molares humanos provenientes del yacimiento del Pleistoceno medio de Pinilla del Valle (Madrid). Trabajos
de Antropología, 19(4): 303. |
| ○ |
Alférez, F. (1987). Paleoambiente y rasgos culturales de los homínidos del Pleistoceno medio de Pinilla del Valle (Madrid).
In: Abstracts of XII International Congress of INQUA, Ottawa: 119. |
| ○ |
Alférez, F. (1990). Mamíferos, Cap. 1 y 2. In: Paleontología, T. 3. Edit. Paraninfo, Madrid: 1-48. |
| ○ |
Alférez, F. & Buitrago, A.M. (1994). Los restos de Sus scrofa (Artiodactyla, Mammalia) del Pleistoceno Medio de Pinilla del Valle (Madrid). In: Comunicaciones de las X Jornadas de Paleontología,
Madrid: 12-13. |
| ○ |
Alférez, F. & Iñigo, C. (1990). Los restos de Dicerorhinus hemitoechus (Perissodactyla; Mammalia) del Pleistoceno medio de Pinilla del Valle (Madrid). Acta Salmanticensia, 68: 25-45. |
| ○ |
Alférez, F. & Molero, G. (1982). Descubrimiento de un fósil humano (Riss-Würm) en Pinilla del Valle (Madrid). Résumés des
Comunications 1er Congrès International de Paléontologie Humaine, Nice: 103-104. |
| ○ |
Alférez, F. & Roldán, B. (1992). Un molar humano Anteneandertal con patología traumática procedente del yacimiento cuaternario
de Pinilla del Valle (Madrid). Munibe, supl. 8, 183-188. |
| ○ |
Alférez, F.; Molero, G.; Maldonado, E.; Bustos, V.; Brea, P. & Buitrago, A.M. (1982). Descubrimiento del primer yacimiento
cuaternario (Riss-Würm) de vertebrados con restos humanos en la provincia de Madrid (Pinilla del Valle). Coloquios de Paleontología,
37: 15-32. |
| ○ |
Alférez, F.; Molero, G.; Maldonado, E.; Brea, P.; Bustos, V.; Buitrago, A.M. & Toni, I. (1983). Paleontología del Cuaternario.
Yacimiento cuaternario de Pinilla del Valle. In: Madrid en sus orígenes (Consejería de Cultura, Deportes y Turismo, ed). Comunidad
de Madrid, 11 pp. |
| ○ |
Alférez, F.; Maldonado, E. & Molero, G. (1985a). El équido del Pleistoceno medio de Pinilla del Valle (Madrid). Equus caballus pinillensis nov. subsp. In: Resúmenes de las Comunicaciones VII Bienal de Real Sociedad Española de Historia Natural, Barcelona: 54. |
| ○ |
Alférez, F.; Molero, G. & Maldonado, E. (1985b). Estudio preliminar del úrsido del yacimiento del Cuaternario medio de Pinilla
del Valle (Madrid). Coloquios de Paleontología, 40: 59-67. |
| ○ |
Altuna, J. (1967). Fauna de mamíferos del yacimiento prehistórico de Marizulo (Urnieta), Guipuzcoa. Munibe, 19: 271-298 |
| ○ |
Altuna, J. (1973). Fauna del yacimiento prehistórico de Los Casares (Guadalajara). Excavaciones arqueológicas en España, 76:
97-116. |
| ○ |
Altuna, J. (1980). Historia de la domesticación animal en el Pais Vasco, desde sus orígenenes hasta la romanización. Munibe,
32: 1-163. |
| ○ |
Altuna, J. & Mariezkurrena, K. (1984). Bases de subsistencia de origen animal en el yacimiento de Ekain. In: El yacimiento
prehistórico de la Cueva de Ekain, Deba, Guipúzcoa (J. Altuna y J. Merino eds). Euskoikaskuntza, B 1: 211-280. |
| ○ |
Álvarez-Lao, D.J.; Arsuaga, J.L.; Baquedano, E. & Pérez-González, A. (2013). Last Interglacial (MIS 5) ungulate assemblage
from the Central Iberian Peninsula: The Camino Cave (Pinilla del Valle, Madrid, Spain). Palaeogeography, Palaeoclimatology,
Palaeoecology, 374: 327-337. https://doi.org/10.1016/j.palaeo.2013.01.025 |
| ○ |
Arribas, A.; Rábano, I. & Gutiérrez-Marco, J.C. (1998). El Patrimonio Paleontológico de la Comunidad de Madrid. In: Patrimonio
Geológico de la Comunidad Autónoma de Madrid (Durán, J.J. ed). Sociedad Geológica de España-Asamblea de Madrid, Madrid: 77-99. |
| ○ |
Arsuaga, J.L.; Baquedano, E.; Pérez-González, A.; Sala, M.T.N.; García, N.; Álvarez-Lao, D.; Laplana, C.; Hughet, R.; Sevilla,
P.; Maldonado, E.; Blain, H.-A.; Quam, R.M.; Ruiz-Zapata, M.B.; Sala, P.; Gil García, M.J.; Uzquiano, P. & Pantoja, A. (2010).
El yacimiento arqueopaleontológico del Pleistoceno Superior de la Cueva del Camino en el Calvero de la Higuera (Pinilla del
Valle, Madrid). Zona Arqueológica, 13: 422-442. |
| ○ |
Arsuaga, J.L.; Baquedano, E.; Pérez-González, A.; Sala, N.; Quam, R.M.; Rodríguez, L.; García, R.; García, N.; Álvarez-Lao,
D.; Laplana, C.; Hughet, R.; Sevilla, P.; Maldonado, E.; Blain, H.-A.; Ruiz-Zapata, M.B.; Sala, P.; Gil-García, M.J.; Uzquiano,
P.; Pantoja, A. & Márquez, B. (2012). Understanding the ancient habitats of the last-interglacial (late MIS 5) Neanderthals
of central Iberia: Paleoenvironmental and taphonomic evidence from the Cueva del Camino (Spain) site. Quaternary International,
275: 55-75. https://doi.org/10.1016/j.quaint.2012.04.019 |
| ○ |
Baquedano, E.; Márquez, B.; Laplana, C.; Gómez, J.; Pérez-González, A. & Arsuaga, J.L. (2015). Creación y musealización del
Parque Arqueológico del Calvero de La Higuera (Pinilla del Valle, Comunidad de Madrid), en el Valle Alto del Lozoya: El Valle
de los Neandertales. Espacio, Tiempo y Forma, Serie I, 8: 155-179. https://doi.org/10.5944/etfi.8.2015.15604 |
| ○ |
Blain, H.-A.; Laplana, C.; Sevilla, P.; Arsuaga, J.L.; Baquedano, A. & Pérez-González, A. (2014). MIS 5/4 transition in a
mountain environment: herpetofaunal assemblages from Cueva del Camino, central Spain. Boreas, 43: 107-120. https://doi.org/10.1111/bor.12024 |
| ○ |
Bochenski, Z. (2007). The Middle Pleistocene avifauna from Pinilla del Valle (Spain). Geobios, 40: 31-37. https://doi.org/10.1016/j.geobios.2005.06.006 |
| ○ |
Bonifay, M.T. (1966). Les artiodactyles non ruminants. II: Le genre Sus. In: Faunes et flores préhistoriques de l’Europe occidentale (Lavocar ed). N. Boubée & Cie, Paris: 225-232. |
| ○ |
Bouchud, J. (1972). Les grand herbivores rissiens des “Abîmes de la Fage” en Corrèze (Cervidés, Bovidés, Capridés, Rupicaprinés,
Suidés et Equidés). Nouvelles Archives du Musée d’Histoire Naturel de Lyon, 10: 35-59. |
| ○ |
Buitrago, A.M. (1992). Estudio de los Artiodáctilos del yacimiento del Pleistoceno medio de Pinilla del Valle (Madrid). Tesis
doctoral, Universidad Complutense de Madrid, 569 pp. |
| ○ |
Carbó, A.; Alférez, F.; Hoster, M.; Carrasco, R. & Domínguez, A. (1990). Prospección geofísica de cavidades naturales en el
yacimiento del Pleistoceno medio de Pinilla del Valle (Madrid). Geociências, Aveiro, 5(1): 1-8. |
| ○ |
Carbonell, E.; Bermúdez de Castro, J.M.; Parés, J.M.; Pérez-González, A.; Cuenca-Bescós, G.; Ollé, A.; Mosquera, M.; Huguet,
R.; Made, J. van der; Rosas, A.; Sala, R.; Vallverdú, J.; García, N.; Granger, D.E.; Martinón-Torres, M.; Rodríguez, X.P.;
Stock, G.M.; Vergès, J.M.; Allué, E.; Burjachs, F.; Cáceres, I.; Canals, A.; Benito, A.; Díez, C.; Lozano, M.; Mateos, A.;
Navazo, M.; Rodríguez, J.; Rosell, J. & Arsuaga, J.L. (2008). The first hominin of Europe. Nature, 452(7186): 465-469. https://doi.org/10.1038/nature06815 |
| ○ |
Carcavilla, L. & Palacio, J. (2010). Geosites: aportación española al patrimonio geológico mundial. Instituto geológico y
Minero de España, Madrid, 231 pp. |
| ○ |
Cohen, K.M.; Finney, S.C.; Gibbard, P.J. & Fan, J.X. (2013). The ICS International Chronostratigraphic Chart. Episodes, 36:
199-204. |
| ○ |
Corvea Porras, J.L.; de Bustamante Gutiérrez, I.; García-Hidalgo, J.F.; Sanz García, J.M. & Mateos Martín, J. (2006). Guía
de puntos de interés didáctico del norte de la Comunidad de Madrid. Universidad de Alcalá de Henares, 120 pp. |
| ○ |
Faure, M. & Guérin, C. (1983). Le Sus scrofa (Mammalia, Artiodactyla, Suidae) du gisement Pleistocène supérieur de Jaurens, à Nespouls, Corrèze, France. Nouvelles Archives
du Musée d’Histoire Naturel de Lyon, 21: 45-63. |
| ○ |
Guérin, C. & Faure, M. (1997). The wild boar (Sus scrofa priscus) from the post-Villafranchian Lower Pleistocene of Untermassfeld. In: Das Pleistozän von Untermassfeld bei Meiningen (Thüringen),
Teil 1 (Khalke, R.-D., ed.). Monographien aus dem Römisch-Germanisches Zentralmuseum Mainz, 40: 375-383. |
| ○ |
Jordá Pardo, F. (2008). Yacimientos de vertebrados del Plioceno y Pleistoceno español. In: Contextos Geológicos españoles.
Una aproximación al patrimonio geológico español de relevancia internacional (A. García-Cortés ed). Instituto Geológico y
Minero de España, Madrid: 171-183. |
| ○ |
Kahlke, H.D. (1965). Die Cerviden-Reste aus den Tonen von Voigtstedt in Thürigen (Das Pleistozän von Voigtstedt). Paläontographica
Abteilung A, 2(2-3): 221-692. |
| ○ |
Kahlke, R.D. (2000). The Early Pleistocene (Epivillafranchian) faunal site of Untermassfeld (Thuringia, central Germany).
Synthesis of new results. Eraul, 92: 123-138. |
| ○ |
Laplana, C. (2014). El registro de micromamíferos en los yacimientos terciarios y cuaternarios de la Comunidad de Madrid.
Actas de las décimas jornadas de Patrimonio Arqueológico en la Comunidad de Madrid: 169-186. |
| ○ |
Laplana, C. & Sevilla García, P. (2016). Nuevos datos sobre los micromamíferos (Roedores, Insectívoros y Quirópteros) del
yacimiento Camino (Pleistoceno Superior, Pinilla del valle, Madrid). In: XXII Jornadas de la Sociedad Española de Paleontología.
Universidad de León: 135-137. |
| ○ |
Laplana, C.; Blain, H.-A.; Sevilla, P.; Arsuaga, J.L.; Baquedano, E. & Pérez-González, A. (2013). Un assemblage de petits
vertébrés hautement diversifié de la fin du MIS5 dans un environnement montagnard au centre de l’Espagne (Cueva del Camino,
Pinilla del Valle, Communauté Autonome de Madrid). Quaternaire, 24: 207-216. https://doi.org/10.4000/quaternaire.6617 |
| ○ |
Made, J. van der (1991). Sexual bimodality in some recent pig populations and application of the findings to study of fossils.
Zeitschrift Säugetierkunde, 56: 81-87. |
| ○ |
Made, J. van der (1996). Listriodontinae (Suidae, Mammalia), their evolution, systematics and distribution in time and space.
Contributions on Tertiary and Quaternary Geology, 33(1-4): 3-254. |
| ○ |
Maldonado, E. (1991). El yacimiento cuaternario de Pinilla del Valle (Madrid). Cuadernos del Instituto de Investigaciones
Científicas y Ecológicas, 38: 9-22. |
| ○ |
Maldonado, E. (1996). Revisión de los équidos del Pleistoceno medio y superior de España. Tesis doctoral, Universidad Complutense
de Madrid, 249 pp. |
| ○ |
Mariezkurrena, K. (1983). Contribución al conocimiento del desarrollo de la dentición y el esqueleto postcraneal de Cervus elaphus. Munibe, 35: 149-202. |
| ○ |
Martínez-Navarro, B.; Ros-Montoya, S.; Espigares, M.P.; Madurell-Malapeira, J. & Palmqvist, P. (2018). Los mamíferos del Plioceno
y Pleistoceno de la Península Ibérica. Revista PH, 94: 206-249. https://doi.org/10.33349/2018.0.4203 |
| ○ |
Muttoni, G.; Scardia, G. & Kent, D.V. (2013). A critique of evidence for human occupation of Europe older than the Jaramillo
subchron (1 Ma): Comment on ‘The oldest human fossil in Europe from Orce (Spain)’ by Toro-Moyano et al. (2013). Journal of
Human Evolution, 65: 746-749. https://doi.org/10.1016/j.jhevol.2013.08.005 |
| ○ |
Muttoni, G.; Scardia, G. & Kent, D.V. (2018). Early hominins in Europe: The Galerian migration hypothesis. Quaternary Science
Reviews, 180: 1-29. https://doi.org/10.1016/j.quascirev.2017.10.031 |
| ○ |
Pérez-González, A.; Karampaglidis, T.; Arsuaga, J.L.; Baquedano, E.; Bárez, S.; Gómez, J.J.; Panera, J.; Márquez, B.; Laplana,
C.; Mosquera, M.; Hughet, R.; Sala, P.; Arriaza, M.C.; Benito, A.; Aracil, E. & Maldonado, E. (2010). Aproximación geomorfológica
a los yacimientos del Pleistoceno Superior del Calvero de la Higuera en el Valle Alto del Lozoya (Sistema Central Español,
Madrid). Zona Arqueológica, 13: 404-419. |
| ○ |
Pérez Ripoll, M. (1977). Los mamíferos del yacimiento musteriense de Cova Negra (Játiva, Valencia). Servicio de Investigación
Prehistórica, Diputación Provincial de Valencia, Serie de trabajos varios, 53: 147. |
| ○ |
Rodríguez, J.; Burjachs, F.; Cuenca-Bescós, G.; García, N.; Made, J. van der; Pérez-González, A.; Blain, H.A.; Expósito, I.;
López-García, J.M.; García-Antón, M.; Allué, E.; Cáceres, I.; Huguet, R.; Mosquera, M.; Ollé, A.; Rosell, J.; Parés, J.M.;
Rodríguez, X.P.; Díez, J.C.; Rofes, J.; Sala, R.; Saladié, P.; Vallverdú, J.; Bennasar, M.L.; Blasco, R.; Carbonell, E. &
Bermúdez de Castro, J.M. (2011). One million years of cultural evolution in a stable environment at Atapuerca (Burgos, Spain).
Quaternary Science Reviews, 30: 1396-1412. https://doi.org/10.1016/j.quascirev.2010.02.021 |
| ○ |
Ruiz-Zapata, B.; Gil-García, M.J. & Uzquiano, P. (2012). Calvero de la Higuera. In: Carrión, J.S. (coord.), Paleoflora y paleovegetación
de la Península Ibérica e Islas Baleares: Plioceno-Cuaternario. Ministerio de Economía y Competitividad, 541-545. |
| ○ |
Sesé, C. & Soto, E. (2000). Vertebrados del Pleistoceno de Madrid. In: Patrimonio Paleontológico de la Comunidad de Madrid
(Morales, J., coord). Arqueología, Paleontología y Etnografía, 6: 216-243. |
| ○ |
Sesé, C. & Soto, E. (2002). Vertebrados del Pleistoceno del Jarama y Manzanares. Zona Arqueológica, 1: 318-337. |
| ○ |
Teichert, M. (1969). Osteometrische Untersuchungen zur Berechnung der Widweristhöle bei vor-und früngeschichtlichen Schewinen,
Habilitationsscrift (Halle Saale) 1966. Kühn-Archiv., 83: 237-292. |
| ○ |
Toñi, I. & Molero, G. (1990). Los roedores (Rodentia, Mammalia) del yacimiento cuaternario de Pinilla del Valle (Madrid).
Acta Salmanticensia, 68: 359-373. |